Download presentation
Presentation is loading. Please wait.

5
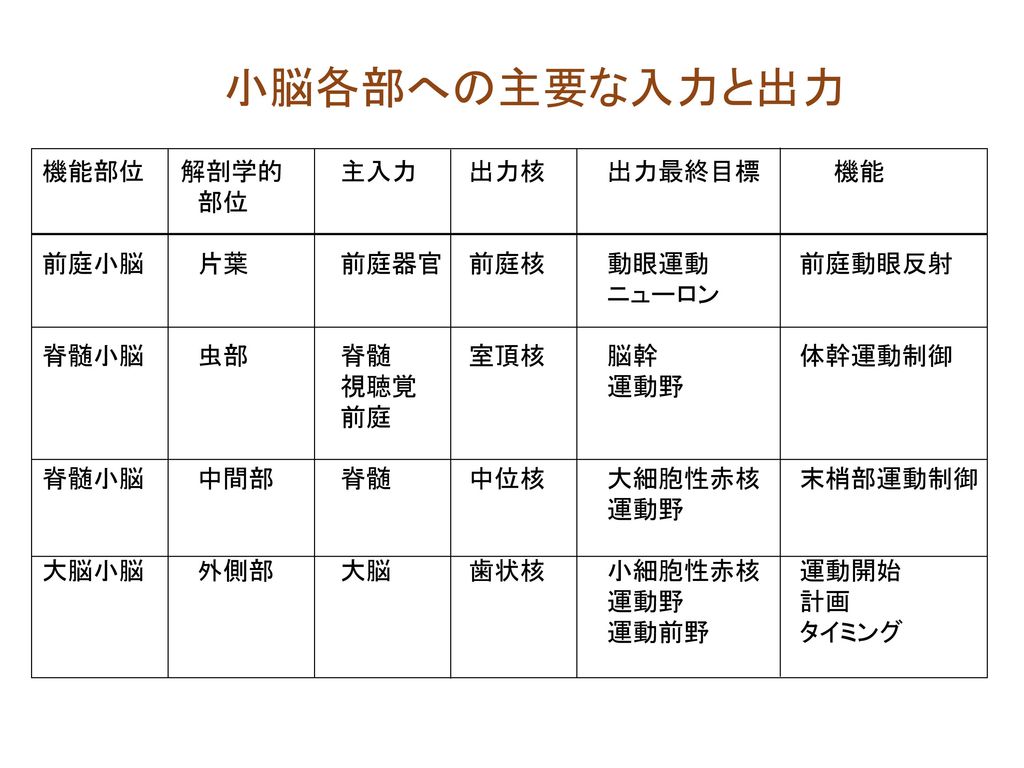
小脳各部への主要な入力と出力 機能部位 解剖学的 主入力 出力核 出力最終目標 機能 部位
機能部位 解剖学的 主入力 出力核 出力最終目標 機能 部位 前庭小脳 片葉 前庭器官 前庭核 動眼運動 前庭動眼反射 ニューロン 脊髄小脳 虫部 脊髄 室頂核 脳幹 体幹運動制御 視聴覚 運動野 前庭 脊髄小脳 中間部 脊髄 中位核 大細胞性赤核 末梢部運動制御 運動野 大脳小脳 外側部 大脳 歯状核 小細胞性赤核 運動開始 運動野 計画 運動前野 タイミング

6
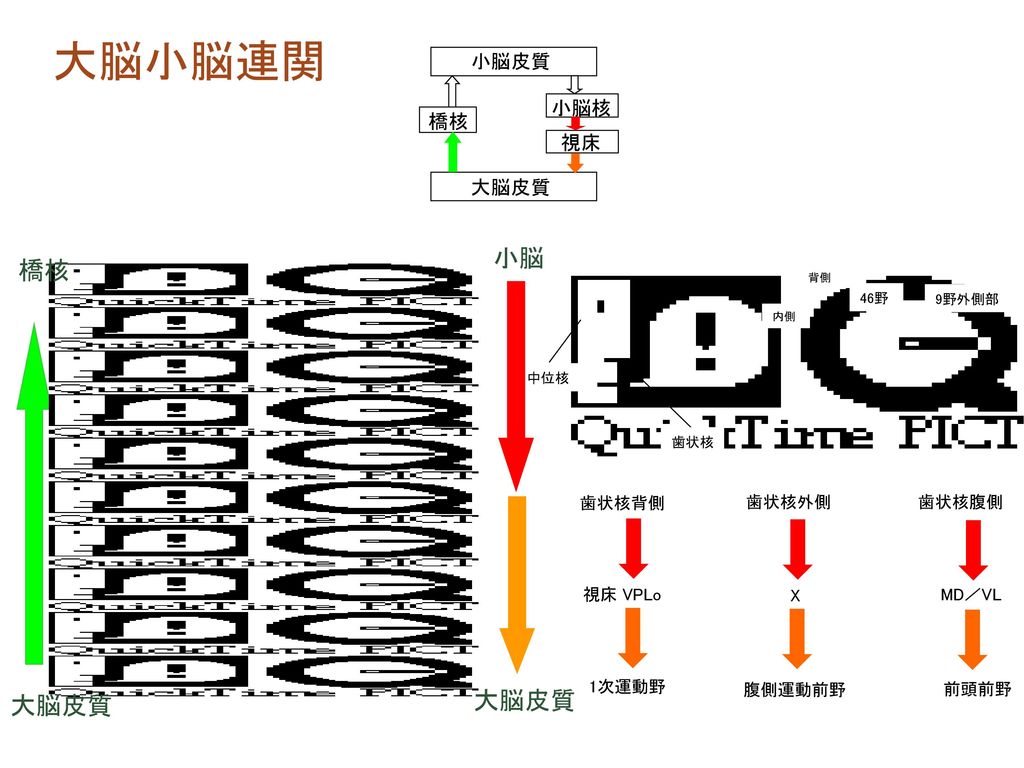
大脳小脳連関 小脳 橋核 大脳皮質 大脳皮質 小脳皮質 小脳核 橋核 視床 大脳皮質 歯状核背側 視床 VPLo 1次運動野 歯状核外側 X
中位核 歯状核 内側 背側 46野 9野外側部 歯状核背側 視床 VPLo 1次運動野 歯状核外側 X 腹側運動前野 歯状核腹側 MD/VL 前頭前野 大脳皮質 大脳皮質

7
小脳の高次認知機能への関与 I. 脳活動計測 II. 患者 III. 解剖
(1) 運動の想像 SPECT Ryding et al. (1993) (2) 名詞からの動詞の連想 PET Raichle et al. (1994) (3) ペグボードパズル fMRI Kim et al. (1994) (4) 複数の形の視覚識別 PET Parsons et al. (1995) (5) 心的回転 PET Parsons et al. (1995) (6) 皮膚感覚による物体認識 fMRI Gao et al. (Bower) (1996) (7) 視覚的注意 fMRI Allen et al. (1997) II. 患者 (1) ハノイの塔 小脳皮質変性症 Grafman et al. (Hallet M.) (1992) (2) 視覚運動認知 小脳皮質変性症 Nawrot & Rizzo (1995) (3) 自閉症患者 小脳サイズ Courchesne et al. (1995) III. 解剖 (1) 46野 HSVI Middleton & Strick (1994) (2) IQ 小脳サイズ Paradiso et al. (1997)
 運動の想像 SPECT Ryding et al. (1993) (2) 名詞からの動詞の連想 PET Raichle et al. (1994) (3) ペグボードパズル fMRI Kim et al. (1994) (4) 複数の形の視覚識別 PET Parsons et al. (1995) (5) 心的回転 PET Parsons et al. (1995) (6) 皮膚感覚による物体認識 fMRI Gao et al. (Bower) (1996) (7) 視覚的注意 fMRI Allen et al. (1997) II. 患者. (1) ハノイの塔 小脳皮質変性症 Grafman et al. (Hallet M.) (1992) (2) 視覚運動認知 小脳皮質変性症 Nawrot & Rizzo (1995) (3) 自閉症患者 小脳サイズ Courchesne et al. (1995) III. 解剖. (1) 46野 HSVI Middleton & Strick (1994) (2) IQ 小脳サイズ Paradiso et al. (1997)")
8
小脳が言葉の連想に 役立っている 名詞を見て理解する(大脳皮質の後ろの視覚に関わる部分)
名詞を聞いて理解する(大脳皮質の横の聴覚に関わる部分) 名詞を見て発音する(大脳皮質の真ん中上と小脳の中心部分) 名詞を見て動詞を連想する(大脳皮質左の言語野と小脳右の外側部分)
 名詞を見て発音する(大脳皮質の真ん中上と小脳の中心部分) 名詞を見て動詞を連想する(大脳皮質左の言語野と小脳右の外側部分)")
10
小脳内部モデル理論 小脳は入力を出力に変換する神経回路の集まり シナプスの伝達効率が変化して異なる変換を学べる
学習は教師がいて、誤差を教えてくれる 脳の外にある、運動のための身体の一部、道具、他人の脳などのまねをする内部モデルを学ぶ

11
ヒトの知性の内部モデル仮説 多数の内部モデルが対になり、多重に、階層的に配置される それが、複雑な行動を計画し、想像し、実行する仕組みである
これは、複雑な運動パターンを認知するため、つまりコミュニケーションにも使われる 言語の機構もこれに違いない

12
高次認知機能の小脳内部モデル仮説 仮説 ・小脳の異なる部位の入出力結合は多様 ・小脳の異なる部位の計算の目的は多様 ・皮質神経回路は一様
・シナプス可塑性は普遍 ・入出力変換の教師有り学習 仮説 (1)系統発生的に新しい部位も古い部位と同様内部モデルを 学習で獲得する (2)内部モデル学習の対象は運動器官に限らない 例:外界の対象物 (道具など)、脳の他の部位、他者 (の脳)
系統発生的に新しい部位も古い部位と同様内部モデルを. 学習で獲得する. (2)内部モデル学習の対象は運動器官に限らない. 例:外界の対象物 (道具など)、脳の他の部位、他者 (の脳)")
13
モザイク:多重の順逆モデル対 (1) 計算論的理由 ・世界はモジュール構造 ・世界は1個の神経回路でモデル化できるほど単純ではない
・異なる学習エポック間での空間的・時間的干渉を減らす ・運動プリミティブ(認知プリミティブ) (2) 支持するデータ ・小脳内の多重スポット:fMRI ・脱適応と再適応は適応より速い ・学習したモジュールの混合 (3) 仮説 多重の逆モデルを学習したりスイッチしたりする最良の 方法は多重の順逆モデル対を学習することである
 (2) 支持するデータ. ・小脳内の多重スポット:fMRI. ・脱適応と再適応は適応より速い. ・学習したモジュールの混合. (3) 仮説. 多重の逆モデルを学習したりスイッチしたりする最良の. 方法は多重の順逆モデル対を学習することである.")
14
l 責任信号 によるモジュールのスイッチング i (1) (1) (4) (3) (2) (3) (4)
責任信号 によるモジュールのスイッチング l i 責任信号:順モデルの予測の相対的良さ (1) ソフトマックス (1) 逆モデル1 順モデル1 運動指令の線形重ね合わせ (4) (3) (2) 逆モデル2 順モデル2 順モデル学習のゲート (3) 逆モデル学習のゲート 逆モデルn 順モデルn (4)
 ソフトマックス. (1) 逆モデル1. 順モデル1. 運動指令の線形重ね合わせ. (4) (3) (2) 逆モデル2. 順モデル2. 順モデル学習のゲート. (3) 逆モデル学習のゲート. 逆モデルn. 順モデルn. (4)")
15
(a)モジュールの学習 (b)スイッチングと制御 M K B 初期状態 対象 責任信号 ステップ 軌 道 1 0.8 0.6 0.4 0.2
-0.2 -0.4 -0.6 0 1 2 3 ×104 軌 道 ステップ 初期状態 対象 9 8 7 6 5 4 3 2 0 1 2 3 4 5 6 7 8 M K B

17
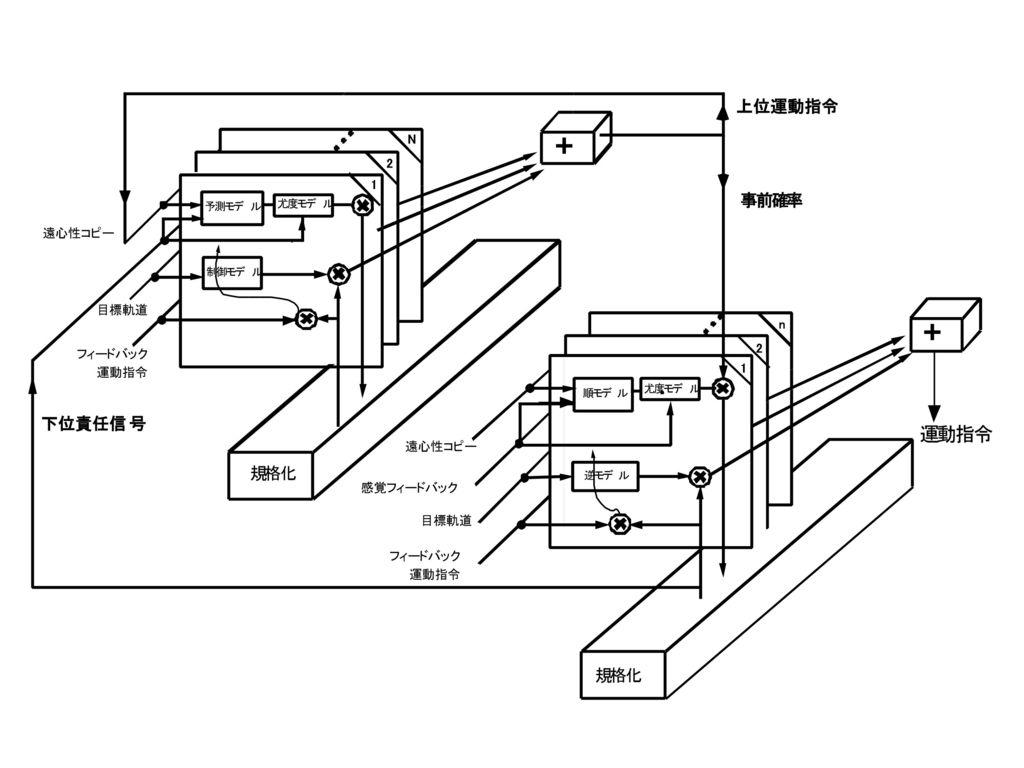
小川の三角形 苔状線維 大脳皮質 苔状線維 遠心性コピー フィードバック 運動指令 目標軌道 マイクロ ゾーン マイクロ ゾーン フィード
フォワード 運動指令 順モデル 運動指令 誤差 逆モデル 感覚フィード バックの予測 感覚 フィード バック 登上線維 登上線維 運動指令 下オリーブ核 歯状核か 大脳皮質 予測誤差 下オリーブ核 歯状核 尤度モデル 尤度 事前確率 登上線維 苔状線維 責任信号 予測器 文脈情報 マイクロ ゾーン 事前確率 責任信号推定器 ソフトマックス 小川の三角形

21
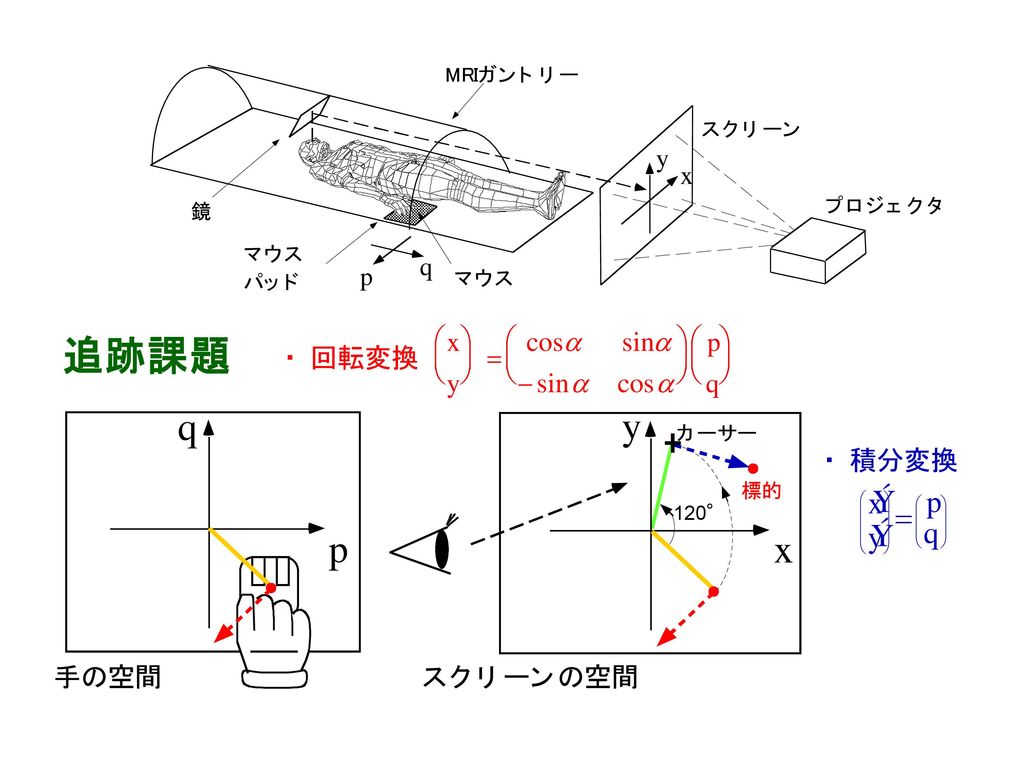
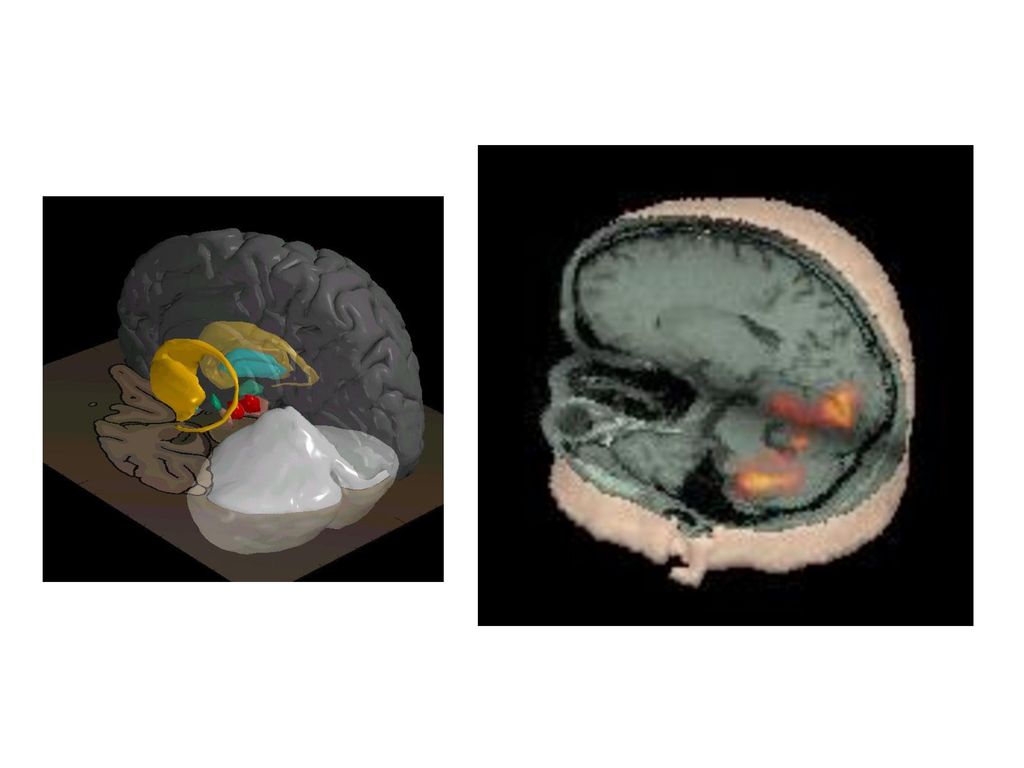
ヒトの脳に 道具の内部モデルがある ヒトの脳の活動を脳を傷つけずに外から計測できる(fMRI, PET、脳波、脳磁図など)
新しい道具を学ぶ時に小脳のたくさんの部分が活動する 特定の道具を使う記憶(内部モデル)は狭い場所に別々にある
は狭い場所に別々にある.")
24
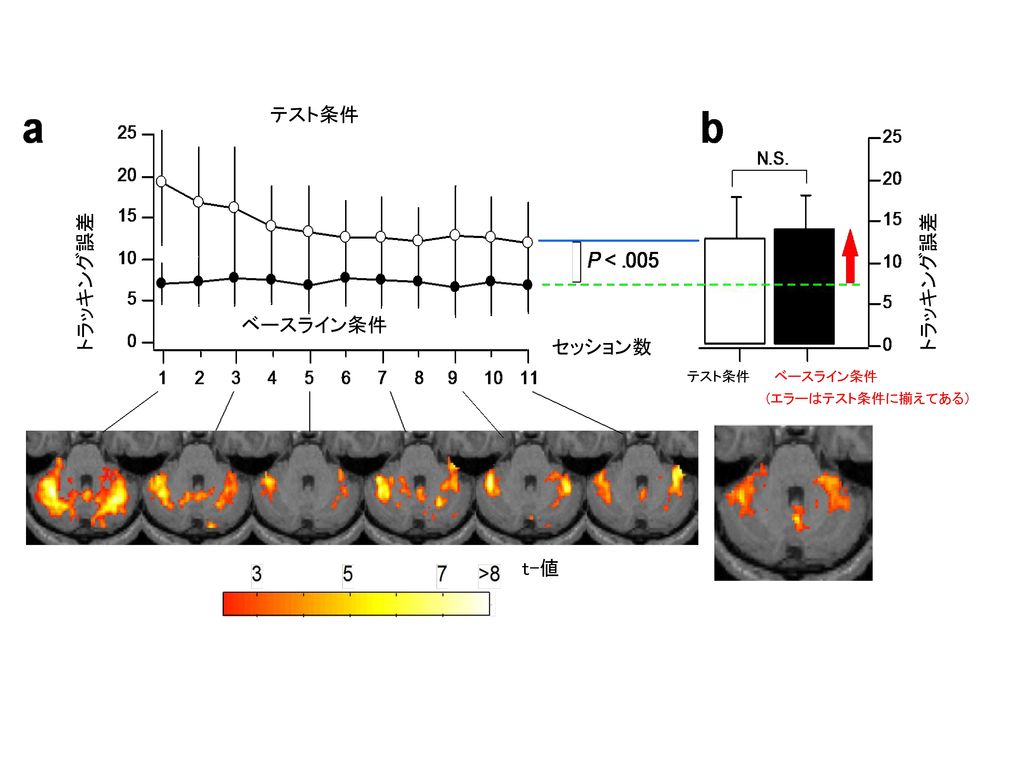
テスト条件 ベースライン条件 トラッキング誤差 セッション数 (エラーはテスト条件に揃えてある) t-値
 t-値")
25
2種類の活動が観察された 1. 学習を誘導する誤差信号 2. 獲得された内部モデル トラッキング誤差と相関の高い領域
1. 学習を誘導する誤差信号 2. 獲得された内部モデル トラッキング誤差と相関の高い領域 学習後,誤差のレベルを揃えた時に タスクと相関の高い領域 と の共通領域 1 . 2 5 1 . 8 . 6 ベースラインからのトラッキング 誤差上昇率(%) ベースラインからのトラッキング 誤差上昇率(%) ベースラインからのMR信号上昇率(%) ベースラインからのMR信号上昇率(%) . 4 . 2 . 1 9 7 5 3 i r エラーを 揃えた セッション エラーを 揃えた セッション セッション数 セッション数
 ベースラインからのトラッキング. 誤差上昇率(%) ベースラインからのMR信号上昇率(%) ベースラインからのMR信号上昇率(%) i. r. エラーを. 揃えた. セッション. エラーを. 揃えた. セッション. セッション数. セッション数.")
26
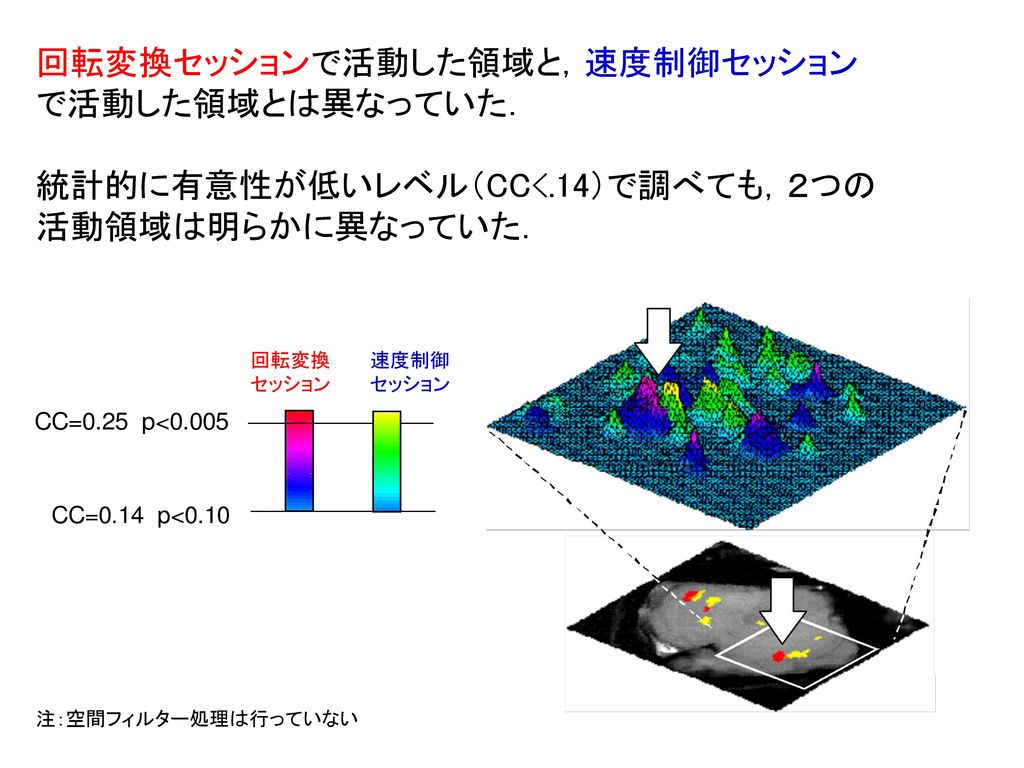
回転変換セッションで活動した領域と,速度制御セッション で活動した領域とは異なっていた.
統計的に有意性が低いレベル(CC<.14)で調べても,2つの 活動領域は明らかに異なっていた. 回転変換 セッション 速度制御 セッション 注:空間フィルター処理は行っていない
で調べても,2つの. 活動領域は明らかに異なっていた. 回転変換. セッション. 速度制御. セッション. 注:空間フィルター処理は行っていない.")
27
B A C 順モデル 対象 手 逆モデル 腕 負荷力 把持力と負荷力 把持力 時間(秒) 把持力 制御器 把持力 負荷力 位置 手の運動
指令 把持力 制御器 順モデル 腕の予測軌道 対象 運動指令の遠心性コピー 手 逆モデル 腕 腕の目標軌道 腕の運動指令

28
把持力調節 A. 物体の移動 B. 負荷腕運動 負荷力 負荷力 粘着テープ で固定 把持力 腕運動 C. 把持力調節 D. 静止状態 把持力

29
腕の運動 把持力調節

30
小脳内の順モデルの異常:分裂病 (Frith, Blakemore, Wolpert)
自分でくすぐってもくすぐったくない 小脳の活動で、体性感覚野の活動が抑制される(Nature Neurosci, 1998; NeuroImage 1999) 分裂病の患者は自分でくすぐっても、くすぐったい 幻聴など多くの症状が、順モデルによる運動指令からの感覚フィードバックの予測の異常と見なせる(Brain Res Rev, 2000)
 分裂病の患者は自分でくすぐっても、くすぐったい. 幻聴など多くの症状が、順モデルによる運動指令からの感覚フィードバックの予測の異常と見なせる(Brain Res Rev, 2000)")
31
制御の妄想の計算モデル 運動制御のモデル 順モデルの異常

32
研究の背景 1. 音節言語は非常に新しい 15万年前に発生 2. Mimesisの理解 ヒトの非言語的コミュニケーション〜300万年前
1. 音節言語は非常に新しい 15万年前に発生 2. Mimesisの理解 ヒトの非言語的コミュニケーション〜300万年前 3. 感覚 - 運動系列学習の大脳・小脳・基底核 ループ神経回路モデル 強化学習+内部モデル+チャンキング 4. 小脳半球腹外側部 道具・他者の脳・脳の一部の内部モデル 5. 埋め込みの神経機構 大脳・小脳コミュニケーションループと小川の三角形 6. 作ることによって知る コミュニケーションのロボットによる再現 音声言語処理スピーチの研究・自然言語処理

34
何故小脳の計算理論がヒト知性の 理解に役立ちうるか? (1) ヒト小脳が高次認知機能に役立っている
(例えばTrends Cogn. Sci ) 小脳の異なる部位の機能はその入出力の違いに対応して実に多様であるが (反射、古典的条件づけ、歩行、随意運動、言語、視覚的注意、心的想像 など)、神経回路は一様であるから、計算原理は共通であると期待される。 小脳の古い部分の計算原理は良く理解されていて運動器官の内部モデルを 学習で獲得することである。新しい部分の計算原理として外界の(道具、 視覚刺激、刺激の時間関係、他人の心、脳の他の部位)内部モデルが考え られる。 小脳半球の後外側部と対応する歯状核腹外側部は、様々の脳部位の中で ヒトで最も著しく増大している。この部位は小川の三角形を介して大脳 前頭葉と回帰的に結合されており、多数の内部モデルを階層的に組み合 わせる神経機構と考えられる。 (2) (3) (4)
 小脳の異なる部位の機能はその入出力の違いに対応して実に多様であるが. (反射、古典的条件づけ、歩行、随意運動、言語、視覚的注意、心的想像. など)、神経回路は一様であるから、計算原理は共通であると期待される。 小脳の古い部分の計算原理は良く理解されていて運動器官の内部モデルを. 学習で獲得することである。新しい部分の計算原理として外界の(道具、 視覚刺激、刺激の時間関係、他人の心、脳の他の部位)内部モデルが考え. られる。 小脳半球の後外側部と対応する歯状核腹外側部は、様々の脳部位の中で. ヒトで最も著しく増大している。この部位は小川の三角形を介して大脳. 前頭葉と回帰的に結合されており、多数の内部モデルを階層的に組み合. わせる神経機構と考えられる。 (2) (3) (4)")
35
仮 説 ヒトの言語システム(ブローカ野、ウェルニッケ野、新小脳 腹外側部)は運動・行動パターンの特に階層的な計画・実行
仮 説 ヒトの言語システム(ブローカ野、ウェルニッケ野、新小脳 腹外側部)は運動・行動パターンの特に階層的な計画・実行 認知・編集(コミュニケーション)に携わる、系統発生的に 古いマカクの相同なシステム(運動前野腹側、上側頭溝、大 脳小脳)から連続的に進化したものである。 (1) ミラーニューロンとPET (Rizzolatti et al., 1992;1996) (2) バイオロジカルモーションの認知 (STS) (Perrett et al., 1989;1990) (3) ヒトとマカクの前頭領野の相同 (Petrides & Pandya, 1994) (4) ヒトの言語コミュニケーションの神経機構の解明は マカクのミメシスを含む非言語的コミュニケーション を研究することで初めて可能となる
は運動・行動パターンの特に階層的な計画・実行. 認知・編集(コミュニケーション)に携わる、系統発生的に. 古いマカクの相同なシステム(運動前野腹側、上側頭溝、大. 脳小脳)から連続的に進化したものである。 (1) ミラーニューロンとPET. (Rizzolatti et al., 1992;1996) (2) バイオロジカルモーションの認知 (STS) (Perrett et al., 1989;1990) (3) ヒトとマカクの前頭領野の相同. (Petrides & Pandya, 1994) (4) ヒトの言語コミュニケーションの神経機構の解明は. マカクのミメシスを含む非言語的コミュニケーション. を研究することで初めて可能となる.")
36
小川の三角形:埋め込み構造、自己回帰 そして階層性のためのハードウェア? 下オリーブ核 下オリーブ核 下オリーブ核

37
コミュニケーションと脳 (1) ブローカ野 文法的複雑さ、複数言語、サインランゲージ、運動の観察と想像 (2) ミラーニューロン
マカクサル腹側運動前野F5はヒトブローカ野(ブロードマン44野)に相同、運動 の観察と実行 (3) 上側頭溝 生物運動パターンの視覚的認知、運動の観察と想像 (4) 小脳外側部 言語、運動の観察と想像、運動前野・上側頭葉と結合 (5) ミメシス ヒト固有の非言語コミュニケーション能力、運動パターンの認知・編集・計画・実行 仮説 ブローカ野・ウェルニッケ野・小脳外側部が運動パターンの認知・編集・計画・ 実行に用いる計算原理はコミュニケーションに普遍である。運動パターンの生成 モデル(逆モデル)と予測モデル(順モデル)を用いた、他者の内部状態の推定 と制御が本質である。
に相同、運動. の観察と実行. (3) 上側頭溝. 生物運動パターンの視覚的認知、運動の観察と想像. (4) 小脳外側部. 言語、運動の観察と想像、運動前野・上側頭葉と結合. (5) ミメシス. ヒト固有の非言語コミュニケーション能力、運動パターンの認知・編集・計画・実行. 仮説. ブローカ野・ウェルニッケ野・小脳外側部が運動パターンの認知・編集・計画・ 実行に用いる計算原理はコミュニケーションに普遍である。運動パターンの生成. モデル(逆モデル)と予測モデル(順モデル)を用いた、他者の内部状態の推定. と制御が本質である。")
38
階層的多重順逆モデル対 (1) シンボル接地問題、チャンキングの問題 を力学系の多重モジュールによる予測と 制御の観点から根源的に解決
い ・・ろは 責予 責予 予 制 予 制 (1) シンボル接地問題、チャンキングの問題 を力学系の多重モジュールによる予測と 制御の観点から根源的に解決 (2) ある階層での責任信号予測器の相互結合 が形成するネットワークは、それ以下の 階層と物理世界が結合して1つの力学系 を形成する (3) 1つ上の階層の順モデルはこの力学系の 状態変化を予測し、逆モデルは文脈情報 と目標軌道を下の階層に与えて、この力 学系を制御する (4) チャンキング(モジュール化)と階層化 は全て双方向の情報の流れの下で自己組 織される (5) モジュールがシンボルに相当し、責任信 号予測器のダイナミクスがシンタックス に相当する。 責予 責予 予 制 予 制 責予 責予 責予 責予 責予 責予 予 制 予 制 予 制 予 制 予 制 予 制 A D A B I II い 1 2 3 4 5 1 2 4 5 3 A B C A C B I II い
 シンボル接地問題、チャンキングの問題. を力学系の多重モジュールによる予測と. 制御の観点から根源的に解決. (2) ある階層での責任信号予測器の相互結合. が形成するネットワークは、それ以下の. 階層と物理世界が結合して1つの力学系. を形成する. (3) 1つ上の階層の順モデルはこの力学系の. 状態変化を予測し、逆モデルは文脈情報. と目標軌道を下の階層に与えて、この力. 学系を制御する. (4) チャンキング(モジュール化)と階層化. は全て双方向の情報の流れの下で自己組. 織される. (5) モジュールがシンボルに相当し、責任信. 号予測器のダイナミクスがシンタックス. に相当する。 責予. 責予. 予 制. 予 制. 責予. 責予. 責予. 責予. 責予. 責予. 予 制. 予 制. 予 制. 予 制. 予 制. 予 制. A. D. A. B. I. II. い A. B. C. A. C. B. I. II. い.")
39
コミュニケーションにおける 階層的な多重の順逆モデル対 (1) 運動パターンの認知から内部状態を推定する
受容した運動パターン→自身の運動パターン →多重モデルが並列的に予測と制御のシミュレーション →最も予測の良い対が推定された内部状態 →隠れた高い階層のダイナミクス(シンボル)の推定 関連:共鳴、共感、心の理論、シミュレーション説、音声知覚の 運動指令説 (2) 運動パターンの提示により内部状態を制御する 他者の (1) の内部モデル及びその逆が必要 →共通の内部モデルの構造を仮定 逆 順 比較
の推定. 関連:共鳴、共感、心の理論、シミュレーション説、音声知覚の. 運動指令説. (2) 運動パターンの提示により内部状態を制御する. 他者の (1) の内部モデル及びその逆が必要. →共通の内部モデルの構造を仮定. 逆. 順. 比較.")
40
1 2 3 4 5 1 2 4 5 3 い ・・ろは 予 制 予 制 予 制 予 制 予 制 予 制 予 制 予 制 予 制 予 制 A D
責予 責予 予 制 予 制 責予 責予 予 制 予 制 責予 責予 責予 責予 責予 責予 予 制 予 制 予 制 予 制 予 制 予 制 A D A B I II い 1 2 3 4 5 1 2 4 5 3 A B C A C B I II い

41
責予 責予 責予 責予 責予 責予 予 制 予 制 予 制 予 制 予 制 予 制 1 2 3 4 5 1 2 4 5 3 l l l l
N l 1 l 1 2 l 1 N 予 制 予 制 予 制 予 制 予 制 予 制 A D A B I II い 1 2 3 4 5 1 2 4 5 3 A B C A C B I II い

42
異なる運動系列のシンボル化 R=0.0009 R=0.001 R=0.002 1-2-1-2-1-3-4-1-3-4-3-4-3-4
sec -π -2π -3π 2π π 1 2 3

43
ACROBOTの振り上げ 生徒 見まね 先生

44
運動パターンの観測から得られた 責任信号予測値による見まね 生徒 見まね 先生 1-2-1-2-3-2-4-2-1-3-2-3-4
7π π π -π -π -π -2π -π -π -π sec sec sec

45
心の理論 (Theory of Mind)
")
46
(Computational Theory of Mind )
心の理論の計算モデル (Computational Theory of Mind ) :) Agent A Agent B (1) A, Bがそれぞれ順に世界を制御、 HMPFIMを獲得 (2) 本質的仮定:2つのHMPFIMは異なるが似ている (3) お互いに相手のモデルを獲得 (4) 上は不可能( 観測不能) (5) imitationのアルゴリズムを用いて他者の を推定、これを用いてtheory of mind学習( (2)) (6) Bootstrap or Annealing Agent B Agent A HMPFIM HMPFIM ~ World
 :) Agent A. Agent B. (1) A, Bがそれぞれ順に世界を制御、 HMPFIMを獲得. (2) 本質的仮定:2つのHMPFIMは異なるが似ている. (3) お互いに相手のモデルを獲得. (4) 上は不可能( 観測不能) (5) imitationのアルゴリズムを用いて他者の を推定、これを用いてtheory of mind学習( (2)) (6) Bootstrap or Annealing. Agent B. Agent A. HMPFIM. HMPFIM. ~ World.")
47
大脳 小脳 大脳基底核 神経回路 双方向+水平結合 一方向 ループ シナプス可塑性 ヘッブ則・反ヘッブ則
異シナプス可塑性 (LTD, LTP, RP) ドーパミンに制御されるヘッブ則 学習形態 教師なし学習 教師あり学習 強化学習 人工的神経回路 連想記憶、コグニトロン パーセプトロン、マダリン アクター・クリティーク モジュール性 領野、コラム 解剖・機能部位、マイクロゾーン 腹背側軸のチャネル、局所モジュール 機能 古 すべて 運動の協調・滑らかさ 行動の選択・安定性 機能 新 すべて すべて すべて 計算原理 統計モデルの学習 力学系の予測と制御モデルの学習 価値関数・報酬関数モデルの学習 フィルター:近距離ガウス分布 水平結合:遠距離コインシデンス 入出力、内部状態 外界のデータ統計に基づく効率的 大脳皮質等で獲得された効率的内部 効率的内部表現と情動に基づく価値 内部表現 表現に基づく因果関係モデル のモデル化 (入出力変換) 計算 低次元への圧縮 因果関係 意味のある因果関係 ひとこと 確率 規則 意味 言語 Lexicon Syntax Semantics 銅谷 (1999)を改変、加筆
 ドーパミンに制御されるヘッブ則. 学習形態. 教師なし学習. 教師あり学習. 強化学習. 人工的神経回路. 連想記憶、コグニトロン. パーセプトロン、マダリン. アクター・クリティーク. モジュール性. 領野、コラム. 解剖・機能部位、マイクロゾーン. 腹背側軸のチャネル、局所モジュール. 機能 古. すべて. 運動の協調・滑らかさ. 行動の選択・安定性. 機能 新. すべて. すべて. すべて. 計算原理. 統計モデルの学習. 力学系の予測と制御モデルの学習. 価値関数・報酬関数モデルの学習. フィルター:近距離ガウス分布. 水平結合:遠距離コインシデンス. 入出力、内部状態. 外界のデータ統計に基づく効率的. 大脳皮質等で獲得された効率的内部. 効率的内部表現と情動に基づく価値. 内部表現. 表現に基づく因果関係モデル. のモデル化. (入出力変換) 計算. 低次元への圧縮. 因果関係. 意味のある因果関係. ひとこと. 確率. 規則. 意味. 言語. Lexicon. Syntax. Semantics. 銅谷 (1999)を改変、加筆.")
48
(More detail: http://www.erato.atr.co.jp/DB/)

49
脳計算モデルによる 6感コミュニケーション (1) 脳の重量 ネアンデルタール大 (2) 数(コミュニケーション) 多数の脳と身体の共同作業
300万年前 ミメシス 共同作業 20-3万年前 言語 高効率 数千年前 記号 時間と空間を超える 数百年前 交通(全身) リアルタイムではない 160年前 電信 記号・リアルタイム 120年前 電話 音声言語・感情 20年前 インターネット 記号→受動的画像・音声 未来 ミメシス+時間+空間+文化 分身コミュニケーション 脳研究+ロボット ●脳のモデル ●見まね制御 ●コミュニケーション 信号の認知 ●コミュニケーション 信号の認知 ●脳のモデル ●見まね制御 ●時間遅れ
 リアルタイムではない. 160年前 電信. 記号・リアルタイム. 120年前 電話. 音声言語・感情. 20年前 インターネット. 記号→受動的画像・音声. 未来 ミメシス+時間+空間+文化 分身コミュニケーション. 脳研究+ロボット. ●脳のモデル. ●見まね制御. ●コミュニケーション. 信号の認知. ●コミュニケーション. 信号の認知. ●脳のモデル. ●見まね制御. ●時間遅れ.")
Similar presentations














.>")








 協調機械システム論 東京大学 人工物工学研究センター 淺間 一.>")
 定常状態の近似 ◎ 反応機構が2ステップを越える ⇒ 数学的な複雑さが相当程度 ◎ 多数のステップを含む反応機構>")