Download presentation
Presentation is loading. Please wait.

1
序 論 生化学とは 生化学はbiochemistryという名で表したように簡単にいえば生命の化学である。ギリシャ語ではbiosは生命という意味である。つまり生化学は化学的理論と技術および物理学、免疫学の原理と方法を応用し、生体における化学構成と化学的変化を研究する学問である。

2
1. 生体分子の構造とその集合状態また構造変化でどのように性質が変わるか?
生化学の主な目的は生命現象を分子レベルで完全に解明することにある。生化学の分野はひろいがその中心課題はつぎの数点にまとめられる。 1. 生体分子の構造とその集合状態また構造変化でどのように性質が変わるか?

3
人体の化学組成はタンパク質,核酸、脂質、糖質,水分、無機物質が主成分である。これらの主成分は厳密な方法に基づいて多種多様な機能を発現する生体構造を構成する。核酸,タンパク質,多糖類は単純生体分子からできているのでバイオポリマーと呼ばれる。これらの生体高分子が本質的には生命の要素を作っている。

4
通常 生体高分子である核酸,タンパク質の構造,機能及び遺伝子構造,発現と制御などの内容を研究する学問は分子生物学と呼ばれている。

5
2. タンパク質の作用機構、酵素触媒の分子機構
3. 遺伝情報の発現機構及び次代への伝達機構 4. 生体分子や生体組織が生成する機構 5. 細胞や生体では無数の生体反応が調和を保つ 機構 6. 細胞の増殖と分化の作用機構

6
生化学と医学 生化学は重要な基礎医学学科の一つである。近年來、生物学、組織学,微生物学,薬理学、病理学などの研究は分子レベルへと発展し,生化学的理論と技術を応用し、それぞれ自分学科の問題を解決するようになる。

7
それと同様に臨床医学では疾患の診断、治療,予防などは生化学の理論と方法に依存するばかりでなく、疾患の原因も分子レベルで解明されるようになった。したがって生化学と医学の間に深い関係がある。医学を志す者は生化学に精通しなければならない。

8
3. 教科書に組み込まれた図解や表を詳しく 読むことは勉強におおいに役立つ。
如何に生化学を学ぶか? 次のいくつかのことを注意しなければならない。 1. 実験を重視するのは大切なことである。 2. 重点をつかんで学ぶこと 3. 教科書に組み込まれた図解や表を詳しく 読むことは勉強におおいに役立つ。 4. 生化学を臨床医学と結びつけて勉強す る。

9
Structure and Function of Protein
第一章 タンパク質の構造と機能 Structure and Function of Protein

10
一、タンパク質とは タンパク質は(protein)多くのアミノ酸同士がぺプチド結合によってつながっている高分子の含窒素化合物である。
多くのアミノ酸同士がぺプチド結合によってつながっている高分子の含窒素化合物である。")
11
二、タンパク質の生物学の重要性 1.構築作用 脂肪組織を除いてタンパク質は細胞や組織を構成する主な有機成分。 2.重要の生物学的機能
1.構築作用 脂肪組織を除いてタンパク質は細胞や組織を構成する主な有機成分。 2.重要の生物学的機能 • 酵素的触媒作用

12
• 輸送と貯蔵 • 協調的運動 • 機械的支持 • 免疫防御 • 神経活動電位の発生と伝達 • 増殖分化の制御 3. 酸化されて エネルギーを供給する

13
タンパク質の分子組成 The Molecular Component of Protein
第一節 タンパク質の分子組成 The Molecular Component of Protein

14
タンパク質の元素組成 単純タンパク質の元素組成は次のようになる。 C、 H、 N(13%-19%)、 O、 S ある物 P、 Fe、 Cu、 Zn、 Mn、 Mo、 Se、 I などを含む。
、 O、 S ある物 P、 Fe、 Cu、 Zn、 Mn、 Mo、 Se、 I などを含む。")
15
その特徴を利用して 生物試料中の窒素測定によって そのタンパク質定量をすることができる。
タンパク質元素組成の特徴 多くのタンパク質の窒素含有量は平均して約16%である。 その特徴を利用して 生物試料中の窒素測定によって そのタンパク質定量をすることができる。

16
タンパク質は約16%の窒素を含んでいるので1グラム窒素が6.25gタンパク質に相当する。つまり
100g試料中のタンパク質の含量(g%) = グラム当たり試料中の窒素g数 ×6.25×100 1/16%
 = グラム当たり試料中の窒素g数 ×6.25×100. 1/16%")
17
一、アミノ酸(amino acid) タンパク質を構成する基本単位
天然界には300を超える種類のアミノ酸が存在するにもかかわらず,そのうち20種類のアミノ酸のみがポリペプチド骨格を構成する単位となっている。

18
L-アミノ酸の共通な構造 R CH3 アラニン H グリシン

19
(一)アミノ酸の分類 1.非極性疎水性アミノ酸 (1∼7) 2.極性中性アミノ酸 (8∼15) 3.酸性アミノ酸 (16∼17)
1.非極性疎水性アミノ酸 (1∼7) 2.極性中性アミノ酸 (8∼15) 3.酸性アミノ酸 (16∼17) 4.塩基性アミノ酸 (18∼20) * 20種類のアミノ酸の英語、略号 及び分類が次のようになる。
 2.極性中性アミノ酸 (8∼15) 3.酸性アミノ酸 (16∼17) 4.塩基性アミノ酸 (18∼20) * 20種類のアミノ酸の英語、略号 及び分類が次のようになる。")
20
イソロイシン isoleucine Ile I 6.02
3文字 略号 1文字 略号 等電点 構造式 日本語 英語 非極性疎水性アミノ酸 グリシン glycine Gly G アラニン alanine Ala A バリン valine Val V ロイシン leucine Leu L イソロイシン isoleucine Ile I フェニルアラニン phenylalanine Phe F プロリン proline Pro P 目 录

21
トリプトファン tryptophan Try W 5.89
2.極性中性アミノ酸 トリプトファン tryptophan Try W セリン serine Ser S チロシン tyrosine Try Y システイン cysteine Cys C メチオニン methionine Met M アスパラギン asparagine Asn N グルタミン glutamine Gln Q トレオニン threonine Thr T 目 录

22
アスパラギン酸 aspartic acid Asp D 2.97
3.酸性アミノ酸 アスパラギン酸 aspartic acid Asp D グルタミン酸 glutamic acid Glu E 4.塩基性アミノ酸 リシン lysine Lys K アルギニン arginine Arg R ヒスチジン histidine His H 目 录

23
特殊のアミノ酸 ‧プロリン (イミノ酸)
")
24
システイン + -HH ジスルフィ ド結合 シスチン

25
中性アミノ酸はR基の構造によりさらに脂肪族アミノ酸(1∼5),芳香族アミノ酸(6,8,10)、水酸基アミノ酸(9,10,13)、含硫アミノ酸(11,12)、イミノ酸,酸アミドアミノ酸に分類される。
,芳香族アミノ酸(6,8,10)、水酸基アミノ酸(9,10,13)、含硫アミノ酸(11,12)、イミノ酸,酸アミドアミノ酸に分類される。")
26
ここではいくつかのことを説明したいと思う。
Ⅰ.上述した20種のアミノ酸は天然タンパク質を構成するもので,それぞれ対応する遺伝暗号がある。いいかえれば 上述した20種のアミノ酸は体内の遺伝暗号で規定されたものである。(標準アミノ酸)
")
27
Ⅱ. 近年來 セレノシステインとよばれる特殊なアミノ酸が発見された。このアミノ酸はタンパク質の構成に関与するばかりでなく対応する遺伝暗号もある。セレノシステインは21番目の標準アミノ酸となる。(R:H-Se-CH2-)。
。.")
28
Ⅲ. 天然タンパク質の中に存在するアミノ酸は上述した20種のアミノ酸だけでない。ほかのアミノ酸もタンパク質の構成成分としてやはり重要なものである。
例えばヒドロキシプロリン、ヒドロキシリシンはコラーゲンを構成する重要なアミノ酸成分である。

29
しかしこのようなアミノ酸は体内では対応する遺伝暗号がない。実際にはこれらはタンパク質が構成されてからあるいは構成されるとき特異的に修飾されて生じたものである。標準アミノ酸と区別するためこれらのアミノ酸を非標準アミノ酸(修飾アミノ酸)と呼ぶ。
と呼ぶ。")
30
Ⅳ. いくつかのアミノ酸は天然タンパク質の構成に関与しないで対応する遺伝暗号もない。これらのアミノ酸はただアミノ酸代謝中間体として一定の役割を果たす。例えば オルニチン、シトルリン、ホモシステインなど(非タンパク質アミノ酸)。
。")
31
(二)アミノ酸の物理化学的性質 1. 両性解離及び等電点 アミノ酸が両性電解質である。その解離程度がそのときの溶液のpHによる。
アミノ酸の物理化学的性質 1. 両性解離及び等電点 アミノ酸が両性電解質である。その解離程度がそのときの溶液のpHによる。")
32
等電点(isoelectric point, pI)
アミノ酸はあるpHの溶液において 分子の正味の荷電が0になる。そのときの溶液のpHをそのアミノ酸の等電点と呼ぶ。

33
+OH- +OH- +H+ +H+ pH>pI pH<pI pH=pI 陽イオン アミノ酸双性イオン 陰イオン

34
2. 紫外吸収 トリプトファン、チロシンの最大の吸収ピークは280nm付近にある。その特徴を利用して生物試料の280nm吸収度の測定によりそのタンパク質定量をすることができる。 芳香族アミノ酸の紫外吸収

35
3. ニンヒドリン反応 (ninhydrin reaction)
アミノ酸とニンヒドリンが反応すると赤紫色の化合物を生成する。この化合物の吸収最大波長が570nmにある。吸収ピーク値がアミノ酸の含量に比例するのでアミノ酸の定量に用いられる。

36
二、ペプチド (一)ペプチド (peptide) タンパク質分子におけるアミノ酸連結様式 * ペプチド結合 (peptide bond)
タンパク質分子におけるアミノ酸連結様式 一つのアミノ酸のα-COOHと他のアミノ酸のα-NH2との間で1分子の水を失って重合し生じた結合をペプチド結合と呼ぶ。

37
+ グリシン グリシン -HOH グリシニルグリシン ペプチド結合

38
* ペプチド(peptide) α-アミノ酸同士はペプチド結合によってつながって形成された化合物をペプチドという。
 α-アミノ酸同士はペプチド結合によってつながって形成された化合物をペプチドという。")
39
* 2個,3個,数個のアミノ酸からなる重合体はそれぞれジペプチド、トリペプチド、オリゴペプチド(oligopeptide)と呼ばれる。通常10個以上のアミノ酸からなるものをポリペプチド(polypeptide)あるいはポリペプチド鎖(polypeptide chain)と呼ぶ。
と呼ばれる。通常10個以上のアミノ酸からなるものをポリペプチド(polypeptide)あるいはポリペプチド鎖(polypeptide chain)と呼ぶ。")
40
* アミノ酸残基(amino acid residue)
アミノ酸がペプチド結合を形成し,タンパク質やペプチドに組み込まれた時、1分子の水を失って 残った部分をアミノ酸残基と呼ぶ。 簡単に言えばタンパク質やペプチド中のアミノ酸単位がアミノ酸残基と呼ばれる。

41
* N末端、C末端 一本のポリペプチド鎖には二つの遊離末端即ちα-NH2とα-COOH基を持っている末端がある。

42
ペプチドでは遊離α-NH2をもつアミノ酸残基をアミノ末端(N末端)といい、左側に書き、H2N-あるいはH-で表す。遊離α-COOH基をもつアミノ酸残基をカルボキシル基(C末端)末端と呼び、右側に書き、-COOH或は-OHで表す。
といい、左側に書き、H2N-あるいはH-で表す。遊離α-COOH基をもつアミノ酸残基をカルボキシル基(C末端)末端と呼び、右側に書き、-COOH或は-OHで表す。")
43
N末端 C末端 ウシ膵臓リボヌクレアーゼ

44
* ジスルフィド結合(disulfide bond)
ポリペプチド鎖の中ではペプチド結合のほかにジスルフィド結合がある。ジスルフィド結合は二つのCys残基が酸化されて形成された結合である。

45
‧鎖内ジスルフィド結合: ポリペプチド鎖内の両部分の間で形成される。 ‧鎖間ジスルフィド結合: 鎖同士の間で形成され,二本ポリペプチド鎖を連結する。

46
1. グルタチオン (glutathione, GSH)
(二)生物活性ペプチド 1. グルタチオン (glutathione, GSH) グルタミン酸 システイン グリシン
生物活性ペプチド. 1. グルタチオン (glutathione, GSH) グルタミン酸. システイン. グリシン.")
47
H2O2 2GSH NADP+ GSHペルオキシダーゼ GSH還元酵素 2H2O GSSG NADPH+H+

48
2.ペプチドホルモン及び神経ペプチド ペプチドホルモンがホルモンの性格を持 ったペプチドの総称である。 例えばチロトロピン放出ホルモン(TRH)が視床下部から分泌されたトリペプチドである。そのN末端はピログルタミン酸で、C末端はPro-NH2である。その生理作用として下垂体前葉から甲状腺刺激ホルモンの分泌を促進する。
が視床下部から分泌されたトリペプチドである。そのN末端はピログルタミン酸で、C末端はPro-NH2である。その生理作用として下垂体前葉から甲状腺刺激ホルモンの分泌を促進する。")
49
ピログルタミン酸 ヒスチジン プロリルアミド チロトロピン放出ホルモン(TRH)
")
50
神経ペプチド (neuropeptide)
神経伝達において情報伝達役割を演ずるペプチドを神経ペプチドとよばれる。 例えば エンケファリン(enkephalin)、エンドルフェン(endorphin)、ダイノルフィン(dynorphin)、サブスタンスP(substance P)、神経ペプチドYなどが知られている。
、エンドルフェン(endorphin)、ダイノルフィン(dynorphin)、サブスタンスP(substance P)、神経ペプチドYなどが知られている。")
51
タンパク質の分子構造 The Molecular Structure of Protein
第二節 タンパク質の分子構造 The Molecular Structure of Protein

52
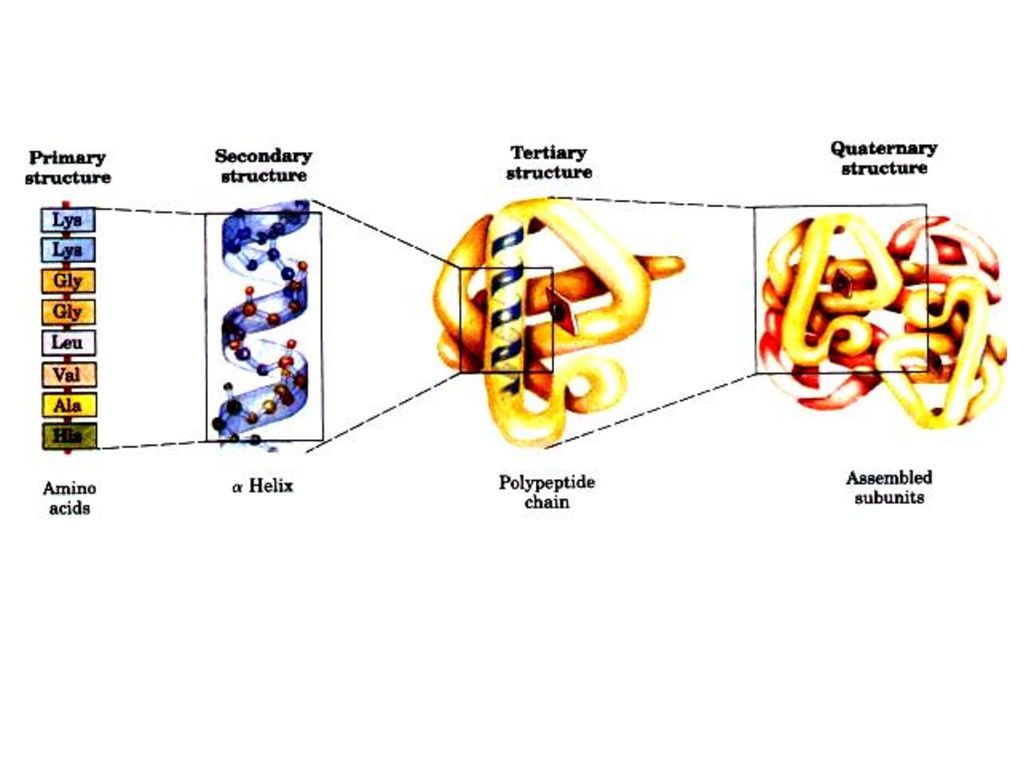
タンパク質はその複雑さによって次の四つの段階に区別する。
• 一次構造 (primary structure) • 二次構造 (secondary structure) • 三次構造 (tertiary structure) • 四次構造 (quaternary structure) 高次構造
 • 二次構造 (secondary structure) • 三次構造 (tertiary structure) • 四次構造 (quaternary structure) 高次構造.")
54
一、タンパク質の一次構造 定義 タンパク質分子中のポリペプチド鎖のアミノ酸配列と もしジスルフィド結合があればその位置をタンパク質の一次構造(primary structure of protein)という。 主な化学結合 : ペプチド結合 あるタンパク質がジスルフィド結合も含ん でいる。

55
タンパク質の一次構造はタンパク質の基本の構造である。これは遺伝子であるDNAの塩基配列によって定められる。
ウシインスリンの一次構造 タンパク質の一次構造はタンパク質の基本の構造である。これは遺伝子であるDNAの塩基配列によって定められる。 目 录

56
タンパク質の一次構造の決定法 Ⅰ タンパク質の材料の調製 2.ポリペプチド鎖の数の決定 3.ジスルフィド結合の切断
1.タンパク質の分離,精製 2.ポリペプチド鎖の数の決定 3.ジスルフィド結合の切断 4.各ポリペプチド鎖の分離,精製 5.ポリペプチド鎖アミノ酸組成の決定

57
Ⅱ ポリペプチド鎖アミノ酸配列の決定 1.ポリペプチド鎖を特定の位置で切断 してアミノ酸配列決定のできる程度の 断片にする。 2.断片の分離,精製 3.各断片のアミノ酸配列の決定 4.ポリペプチド鎖を上と異なる位置で切 断し, アミノ酸配列を決定する。

58
全体の配列を決定する。 2.ジスルフィド結合(鎖内と鎖間)の決定 Ⅲ ポリペプチド鎖全配列の決定 (ペプチド断片の配列順序の決定)
Ⅲ ポリペプチド鎖全配列の決定 (ペプチド断片の配列順序の決定) 1.切断点の異なる断片の配列を比較して 全体の配列を決定する。 2.ジスルフィド結合(鎖内と鎖間)の決定
 1.切断点の異なる断片の配列を比較して. 全体の配列を決定する。 2.ジスルフィド結合(鎖内と鎖間)の決定.")
59
アミノ酸組成の決定 ポリペプチド鎖の加水分解は酸、アルカリによる化学方法と酵素方法があり,それぞれ欠点がある。

60
酸分解では ポリペプチドを6mol/L HClに溶かして真空にした封管の中で ℃ 時間加熱する。しかし Trpは相当破壊され, GlnとAsnはGluとAsp ++NH4に変わるので求めるのはGlx(Glu+Gln)とAsx(Asp+Asn)及び+NH4である。
とAsx(Asp+Asn)及び+NH4である。")
61
酵素による分解 ポリペプチドを完全に酵素で分解するにはいくつかの酵素を混ぜて行う。
アルカリ分解では2-4mol/L NaOHの中で100℃ 4-8時間行う。しかし Cys、Ser、Thr、Argが破壊され、またアミノ酸の一部分は脱アミノ、ラセミを起こす。したがってアルカリ分解は主としてTrpの定量に用いられる。 酵素による分解 ポリペプチドを完全に酵素で分解するにはいくつかの酵素を混ぜて行う。

62
よく使われるのはEdman分解法である。 Edman分解法の基本原理
アミノ酸配列の決定 N末端分析: よく使われるのはEdman分解法である。 Edman分解法の基本原理 1.フェニルイソチオシアネート(phenyl isothiocyanate, PITC, Edman試薬)を弱アルカリ性溶液でタンパク質のN末端アミノ基に作用させ、付加する。
を弱アルカリ性溶液でタンパク質のN末端アミノ基に作用させ、付加する。")
63
2.この付加体は無水フッ化水素酸(FH)をはたらかせるとN末端残基がフェニルチオヒダントイン誘導体として離れるが他のペプチド結合は加水分解されない。つまりN末端アミノ酸残基だけとれ,残りのペプチドはもとのままである。
をはたらかせるとN末端残基がフェニルチオヒダントイン誘導体として離れるが他のペプチド結合は加水分解されない。つまりN末端アミノ酸残基だけとれ,残りのペプチドはもとのままである。")
64
3.フェニルチオヒダントインアミノ酸は高圧液体クロマトグラフィ、電気詠動などで標準物質と比較して同定する。

65
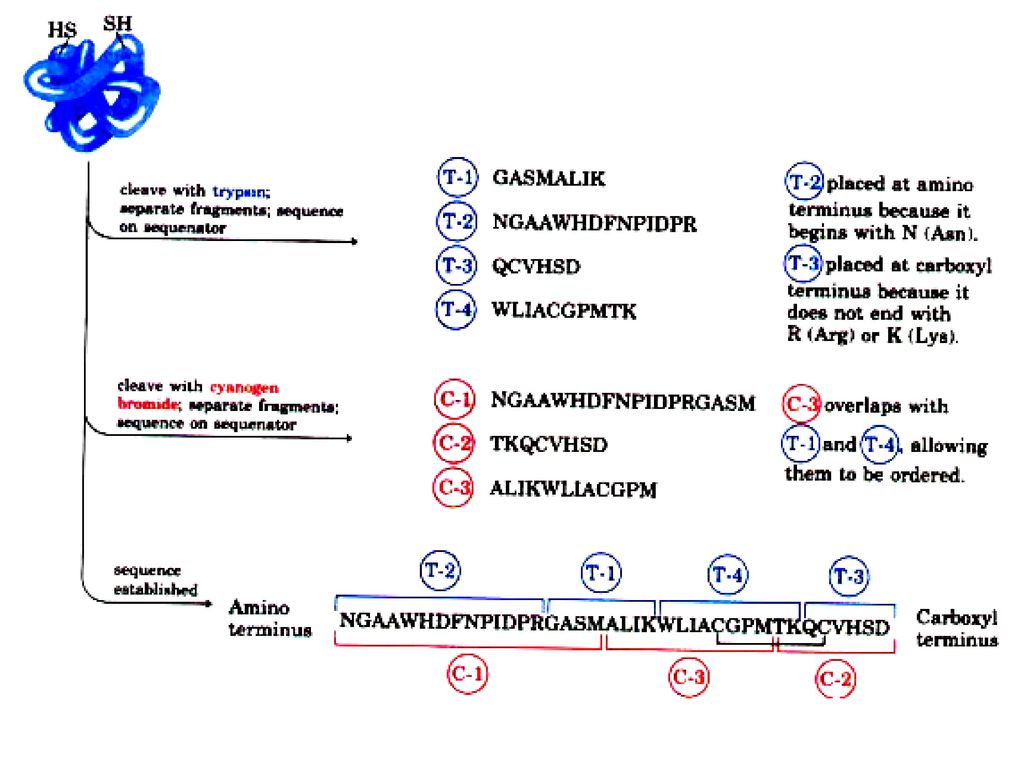
ペプチド結合の特異的切断 40-80残基を超えるポリペプチドはそのままでは配列決定できない。そこで化学的または酵素的に適当な長さに切断する。

66
The speficity of some important methods
for fragmenting polypeptide chains Treatment Cleavage points Trypsin Lys,Arg (C) Chymotripsin Phe,Trp,Tyr (C) V8 protease Asp,Glu (C) Pepsin Phe,Trp,Tyr (N) Cyanogen bromide Met (C)
 Chymotripsin. Phe,Trp,Tyr (C) V8 protease. Asp,Glu (C) Pepsin. Phe,Trp,Tyr (N) Cyanogen bromide. Met (C)")
67
ペプチド断片の配列順序 ペプチド断片のアミノ酸配列が決まったら今度断片の配列順序を調べる。これは最初の断片とは違う位置で切断した異なるペプチド組みの配列を調べ,重複部分を明らかにすればよい。重複部分が十分長ければペプチドの順番がわかる。

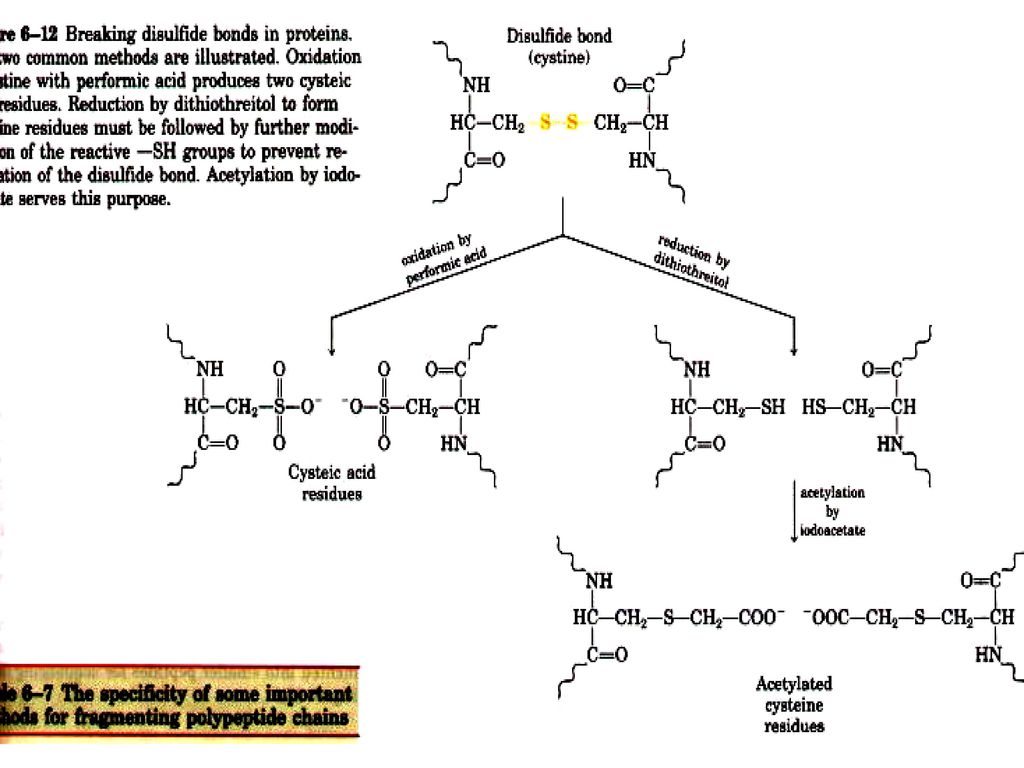
70
ジスルフィド結合の切断 ジスルフィド結合は過ギ酸酸化か、β‒メルカプトエタノール(β‒ME)で還元で切断できる。
で還元で切断できる。")
72
二.タンパク質の二次構造 定義: タンパク質の二次構造(secondary structure of protein)とはポリペプチド主鎖の局部的な空間配置をいう。アミノ酸の側鎖の立体構造に触れない。
とはポリペプチド主鎖の局部的な空間配置をいう。アミノ酸の側鎖の立体構造に触れない。")
73
タンパク質分子では規則的にくり返り部分を主鎖と呼ぶ。
つまり 主鎖はα‐炭素原子とペプチド結合からなっている。 …CαCONHCαCONH… 主な化学結合: 水素結合

74
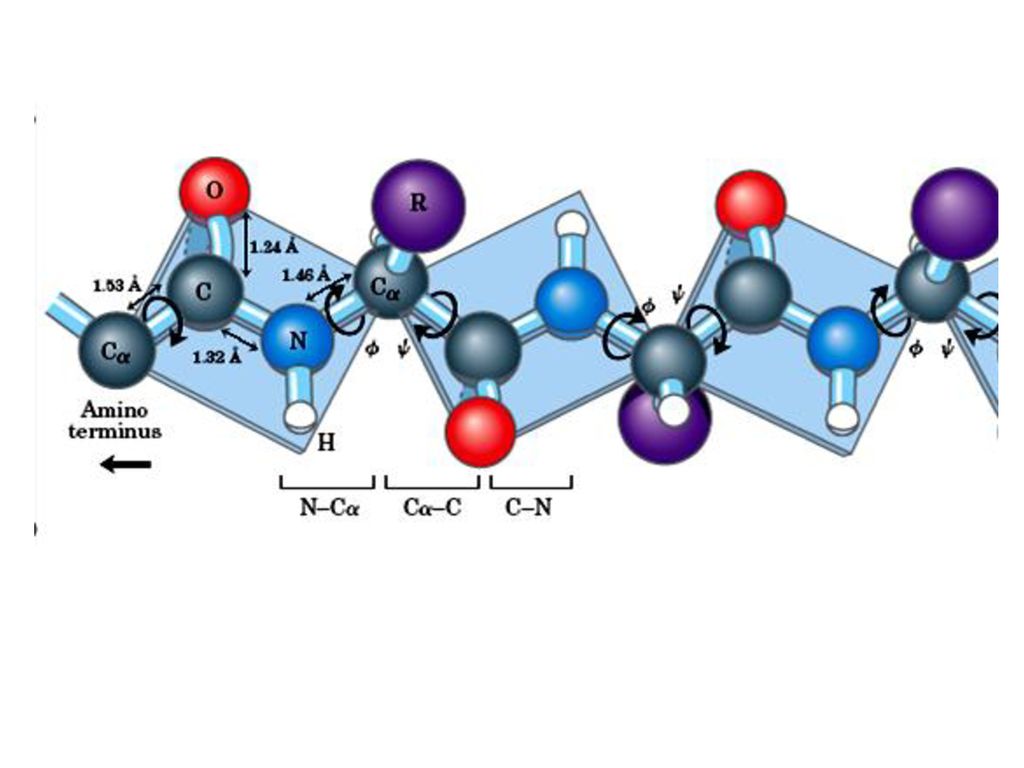
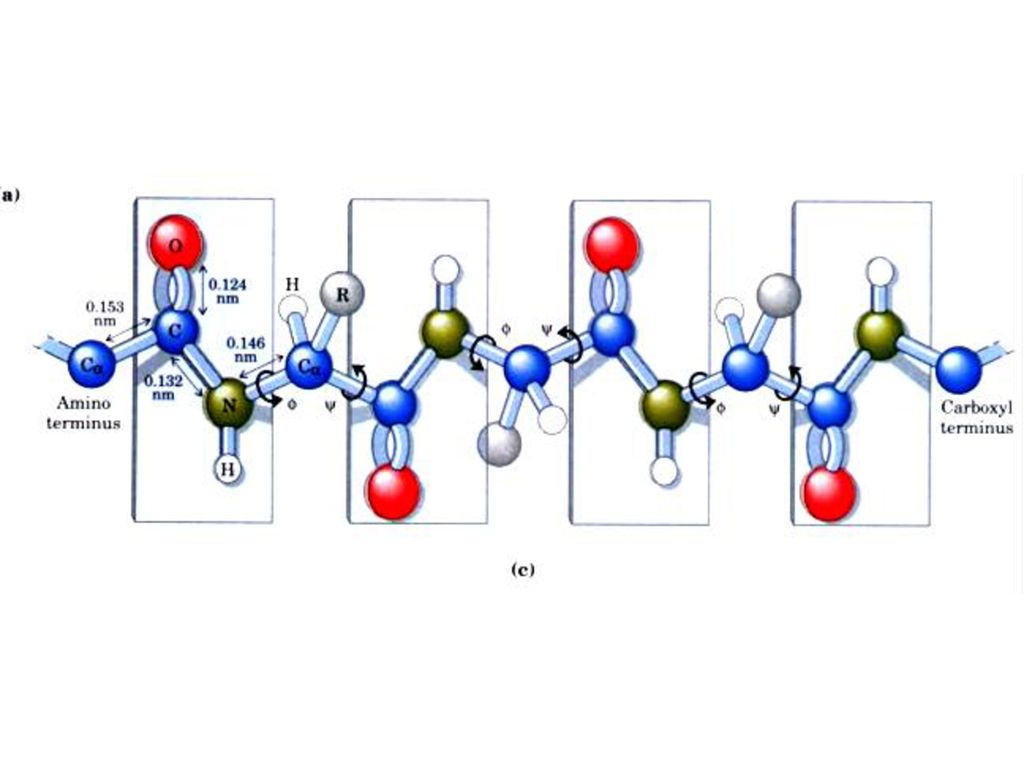
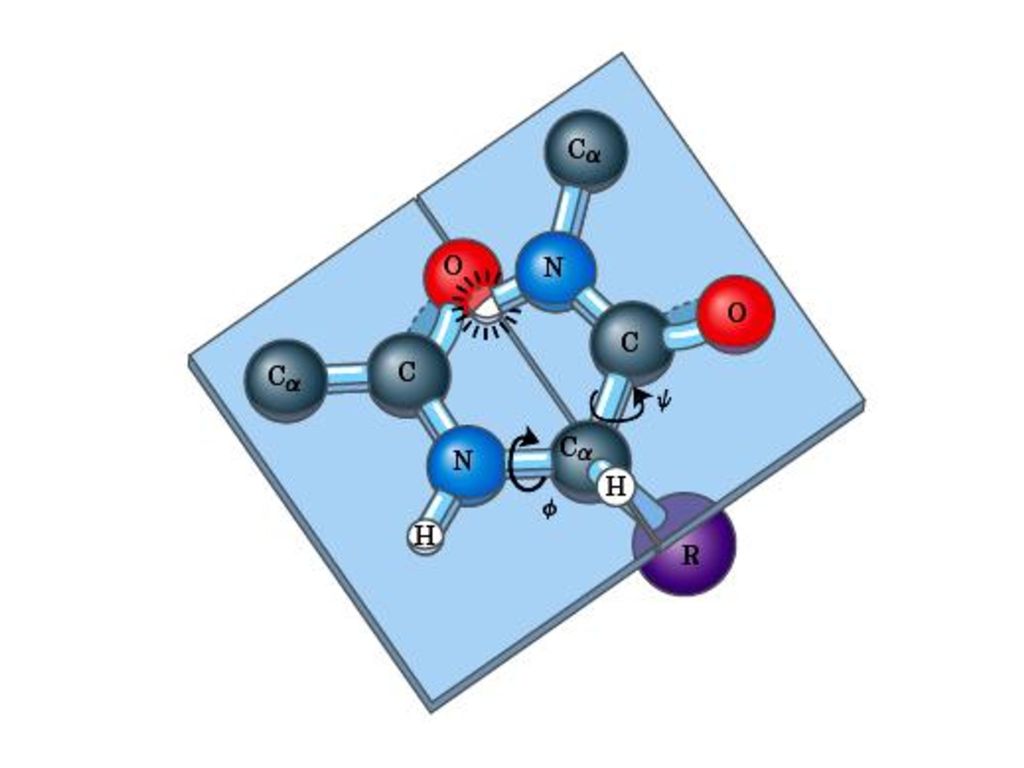
(一)ペプチド結合平面 ポ‒リングとコ‒リ‒はポリペプチド鎖の中でアミノ残基やペプチド結合がどのような制約を受けるかを調べるためにアミノ酸とペプチドの構造のX線結晶解析を行った。その結果 ペプチド結合はしっかりした平面構造をとることがわかった。ペプチド結合平面はタンパク質ポリペプチド主鎖の基本単位である。

75
1.ペプチド結合の長さは0. 132nmであるがそれはC-N単結合(0. 149 nm)とC=N二重結合(0
1.ペプチド結合の長さは0.132nmであるがそれはC-N単結合(0.149 nm)とC=N二重結合(0.127nm)の中間にある。したがってペプチド結合は部分的な二重結合の性質をもって自由に回転できない。ペプチド炭素と窒素周りの結合の角度の和はいずれも360°である。そうすれば図のようにすべての六つの原子は同一面に置かれペプチド平面を形成する。
とC=N二重結合(0.127nm)の中間にある。したがってペプチド結合は部分的な二重結合の性質をもって自由に回転できない。ペプチド炭素と窒素周りの結合の角度の和はいずれも360°である。そうすれば図のようにすべての六つの原子は同一面に置かれペプチド平面を形成する。")
78
2.同一平面にある四つの原子はふつう トランス(trans)に位置している。
3.ペプチド結合のどちらかの側にある結合はある程度自由回転できる。 φ:主鎖の窒素とCα原子の間の結合における回転の量 ψ:Cαとカルボニル炭素の間の結合における回転の量

80
二次構造の主なコンホメーションの様式 α‒ヘリックス ( -helix ) ‒プリ‒ツ•シ‒ト ( -pleated sheet ) ‒タ‒ン ( -turn ) ランダムコイル ( random coil )
 ‒プリ‒ツ•シ‒ト ( -pleated sheet ) ‒タ‒ン ( -turn ) ランダムコイル ( random coil )")
81
(二)α‒ヘリックス(α‒ helix) 目 录
α‒ヘリックス(α‒ helix) 目 录")
82
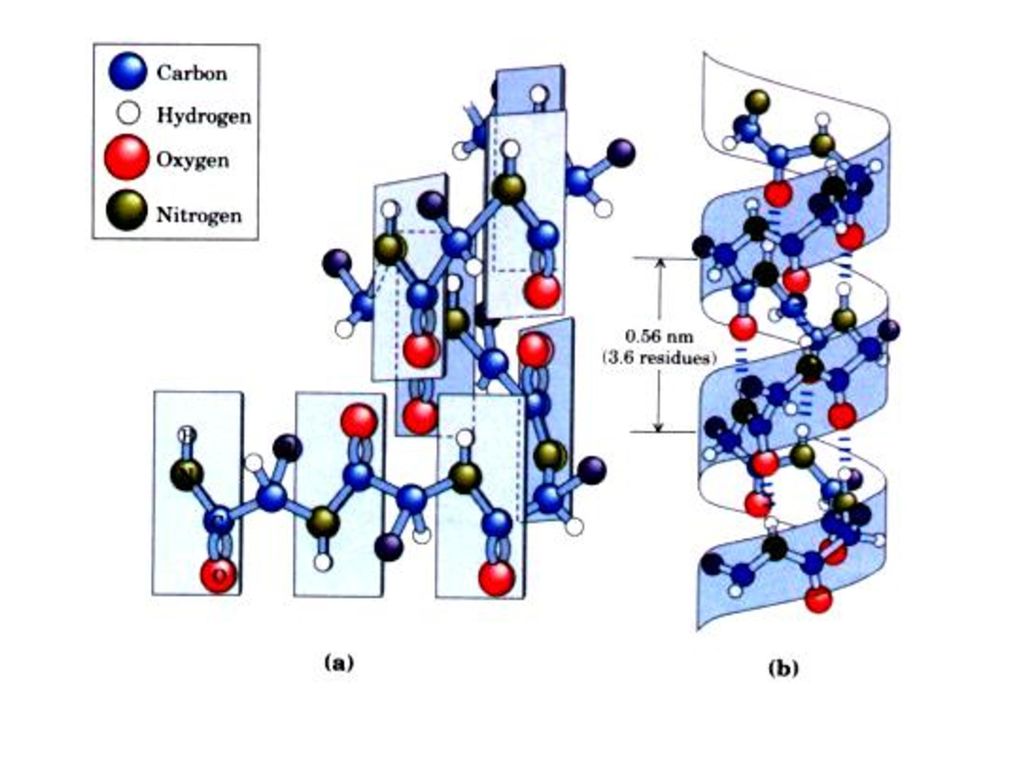
その大切な特徴は次のようになる。 1.右巻きのらせん(時計回り)を形成する。
を形成する。")
83
2.らせんの一回りごとに3. 6個のアミノ酸残基がある。各残基が次の残基とらせんの軸に沿って0
2.らせんの一回りごとに3.6個のアミノ酸残基がある。各残基が次の残基とらせんの軸に沿って0.15nm離れて100°回転している。α‒ヘリックスのピッチは0.54nmである。

84
3.水素結合によって安定化される。 α‒ヘリックスは主鎖のペプチド結合の窒素にくっついた水素原子とカルボニル酸素とで作られた水素結合によって安定化される。(鎖内水素結合)
")
85
水素結合の方向は大体らせん軸に平行している。
各アミノ酸カルボニル基(C=O)は線状配列上4残基先のアミノ酸のNHと水素結合している。この間には13個の原子を含んでいる。 水素結合の方向は大体らせん軸に平行している。 各アミノ酸の側鎖はらせんの外方に突出する。
は線状配列上4残基先のアミノ酸のNHと水素結合している。この間には13個の原子を含んでいる。 水素結合の方向は大体らせん軸に平行している。 各アミノ酸の側鎖はらせんの外方に突出する。")
88
(三)‒プリ‒ツ•シ‒ト ( ‒pleated sheed)
‒プリ‒ツ•シ‒ト ( ‒pleated sheed)")
89
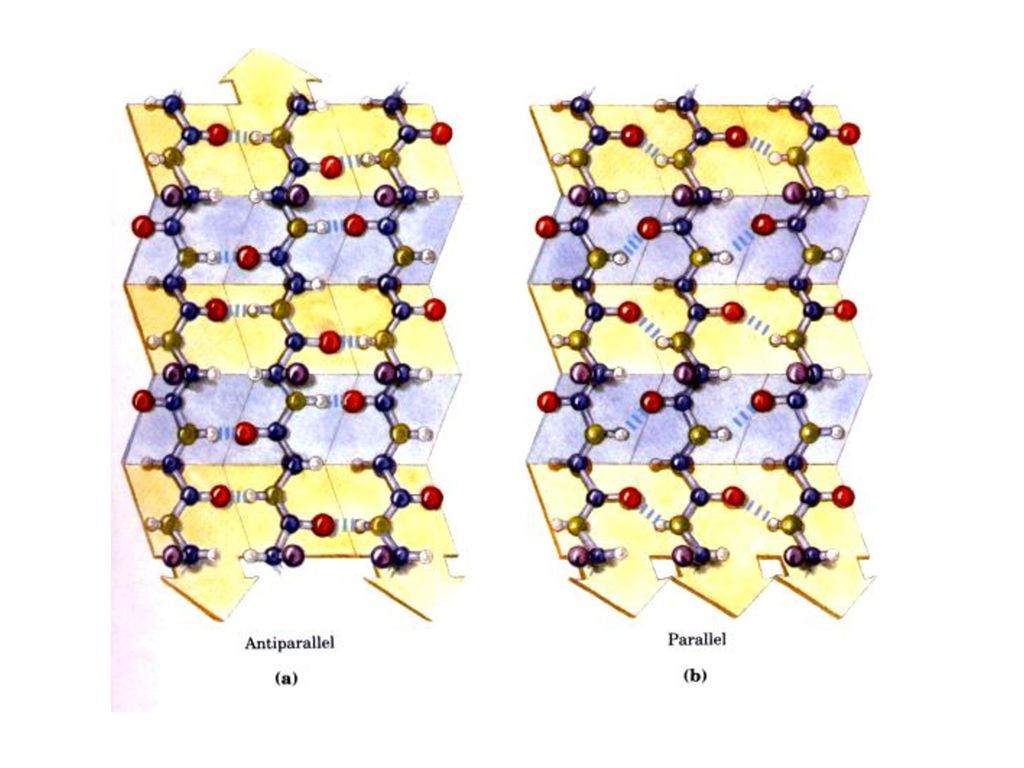
β‒プリ‒ツ•シ‒トはかん状でなく、薄板であるという点でα‒ヘリックスとは顕著に異なっている。
1.完全に延び切ってひだ状(ジクザク状) になる。
 になる。")
90
2.水素結合によって安定化される。 β‒プリ‒ツ•シ‒トは並行する2本以上ポリペプチド鎖あるいは同じペプチド鎖内の両部分からなっている。

91
側鎖は交互に上下に出る。同じ向きに出る周期は0.70nmである。
β‒プリ‒ツ•シ‒トは異なるポリペプチド鎖中のNHとCO基の間の水素結合によって安定化される。(鎖間水素結合) 側鎖は交互に上下に出る。同じ向きに出る周期は0.70nmである。 3.平行β‒プリ‒ツ•シ‒トと逆平行β‒プリ‒ツ •シ‒トに分けられる。
 側鎖は交互に上下に出る。同じ向きに出る周期は0.70nmである。 3.平行β‒プリ‒ツ•シ‒トと逆平行β‒プリ‒ツ •シ‒トに分けられる。")
93
(四)‒タ‒ンとランダムコイル ‒タ‒ン: ポリペプチド鎖がほとんど180°折り返す特異な構造を指す
‒タ‒ンとランダムコイル ‒タ‒ン: ポリペプチド鎖がほとんど180°折り返す特異な構造を指す")
94
ポリペプチド鎖のうち規則性が明確に定められない部分を指す。
よく見られるβ‒タ‒ンは4個のアミノ酸残基からなる。これは球状タンパク質を形成するには必須の領域である。 ランダムコイル(random coil) ポリペプチド鎖のうち規則性が明確に定められない部分を指す。
 ポリペプチド鎖のうち規則性が明確に定められない部分を指す。")
95
通常一つのモチーフは特徴性のアミノ酸配列順序があり、特殊な機能を発揮する。
(五) モチーフ (motif) 多くのタンパク質分子の中で2個または3個の二次構造を持っているペプチド鎖部分が空間的に接近しあい,特殊な機能を持つ空間構造を形成する。そのような空間構造をモチーフという。 通常一つのモチーフは特徴性のアミノ酸配列順序があり、特殊な機能を発揮する。
 モチーフ (motif) 多くのタンパク質分子の中で2個または3個の二次構造を持っているペプチド鎖部分が空間的に接近しあい,特殊な機能を持つ空間構造を形成する。そのような空間構造をモチーフという。 通常一つのモチーフは特徴性のアミノ酸配列順序があり、特殊な機能を発揮する。")
96
Ca2+結合タンパク質に見られるCa2+結合モチーフ 亜鉛フィンガー構造
α‒ヘリックス α‒ヘリックス ループ Ca2+結合タンパク質に見られるCa2+結合モチーフ 亜鉛フィンガー構造

97
(六)アミノ酸のR基がα‒ヘリックスの形成に影響を及ぼす。
同じ電荷を持つアミノ酸が集まった所で はα‒ヘリックスの形成によくない(静 電気的斥力)。
。")
98
大きなR基を持ったアミノ酸(例えば Asn, Ile)はα‒ヘリックスの形成を妨げる(立体障害)
Proはα‒ヘリックスを中断させる。 (側鎖が環状でイミノ窒素には水素原子がない)
")
99
三.タンパク質の三次構造 (一)三次構造(tertiary structure of protein)
定義 タンパク質のポリペプチド鎖はα‒ヘリックスやβ‒フリツ•シ‒ト構造といった二次構造が不規則構造部分を介してさらに折りたたみ,折曲がって形成された一定の規律をもつ三次元構造をタンパク質の三次構造という。

100
ミオグロビン (Myoglobin, Mb) N 端 C端
 N 端 C端")
101
塩結合 疎水結合 主な化学結合: ファンデルワ‒ルス力
簡単に言えば三次構造は主鎖の空間的コンホメ‒ションと側鎖原子の空間的配座及び相互関係を含むポリペプチド全体の三次元構造である。 水素結合 塩結合 疎水結合 ファンデルワ‒ルス力 主な化学結合:

102
それはジスルフィド結合、水素結合、塩結合、疎水結合、ファンデルワ‒ルス力を含む。そのうち重要なものは疎水結合である。
タンパク質の三次構造を維持する結合 三次構造を安定化する因子は側鎖間の相互作用である。 それはジスルフィド結合、水素結合、塩結合、疎水結合、ファンデルワ‒ルス力を含む。そのうち重要なものは疎水結合である。

103
水素結合(hydrogen bond): 水素原子を介して行われる非共有結合の一種である。一般的にOH,NHなどの陽子供与体と電気陰性度の高いCO基などの陽子受容体との間に働き、X-H…Yのよに表される。
: 水素原子を介して行われる非共有結合の一種である。一般的にOH,NHなどの陽子供与体と電気陰性度の高いCO基などの陽子受容体との間に働き、X-H…Yのよに表される。")
104
塩結合(salt linkage) : 異なる電荷をもつ解離基がファンデルワ‒ルス半径まで接近し、クロン力によるイオン対で形成された結合である。
 : 異なる電荷をもつ解離基がファンデルワ‒ルス半径まで接近し、クロン力によるイオン対で形成された結合である。")
105
疎水結合(hydrophobic bond):
水と親和性の少ない非極性基が水溶液の中で互いに集まろうと相互作用である。 球状タンパク質の立体構造では疎水基が分子の内部に、親水基が表面に位置する。

106
ファンデルワ‒ルス力 (van der Waals attraction) 非特異的な引力である。どんな二つ原子でも3-4Å離れたところへ来ると働き出す。
 非特異的な引力である。どんな二つ原子でも3-4Å離れたところへ来ると働き出す。")
107
a.水素結合 b.塩結合 c.疎水結合 目 录

108
ドメインはタンパク質の機能に関係があることが多い。
(二)ドメイン (domain) 構造の面から分子量の大きなタンパク質とくに球状タンパク質にはその立体構造がいくつかの構造単位にわかれることがある。これらの構造的に独立した単位をドメインと呼ぶ。 ドメインはタンパク質の機能に関係があることが多い。
ドメイン (domain) 構造の面から分子量の大きなタンパク質とくに球状タンパク質にはその立体構造がいくつかの構造単位にわかれることがある。これらの構造的に独立した単位をドメインと呼ぶ。 ドメインはタンパク質の機能に関係があることが多い。")
109
フィブロネクチン(fibronectin) に 見られるドメイン
見られるドメイン フィブリン ヘパリン アクチン コラーゲン フィブリン 細胞 フィブリン ヘパリン

110
(三)分子シャペロン (chaperon)
それはタンパク質の正しい空間コンホメ‒ションの形成に関与する一種のタンパク質である。

111
誤った集中の発生を防ぐ ポリペプチド鎖を正しくホールデイングさせる。 タンパク質分子にあるジスルフィド結合の正しい形成に対して重要な役割を果たす。

112
四.タンパク質の四次構造 定義 二本またはそれ以上三次構造を持っているポリペプチド鎖が非共有結合で会合し,特異的な空間配座をとる構造を四次構造(quaternary structure)という。
という。")
113
このようなタンパク質をオリゴマー(oligomer)、ポリマー (polymer) と呼ぶ。
三次構造を持っている各ポリペプチド鎖をプロトマー (protomer)、モノマー(monomer)、サブユニット (subunit) と呼ぶ。 このようなタンパク質をオリゴマー(oligomer)、ポリマー (polymer) と呼ぶ。
、モノマー(monomer)、サブユニット (subunit) と呼ぶ。 このようなタンパク質をオリゴマー(oligomer)、ポリマー (polymer) と呼ぶ。")
114
多くのオリゴマータンパク質は2個あるいは4個のサブユニットを含んでおり、それぞれ二量体(ダイマー, dimmer)、四量体(テトラマー, tetramer) と呼ばれる。
サブユニット間の非共有結合は主に塩結合、水素結合である。

115
サブユニットの会合と解離は可逆的である。モノマーとポリマーの相互変換はタンパク質の機能に関係がある。
正常成人ヘモグロビン(Hb)では2種類のポリペプチド鎖が2個ずつ(α2β2)からなる。したがってHbは四量体構造のタンパク質といえる。
では2種類のポリペプチド鎖が2個ずつ(α2β2)からなる。したがってHbは四量体構造のタンパク質といえる。")
116
ヘモグロビン(Hb)の四次構造
の四次構造")
117
四次構造を持っているタンパク質には同じサブユニットからなるもの(ホモポリマー,homopolymer)もあれば、異なるもの(ヘテロポリマー, heteropolymer)もある。
もあれば、異なるもの(ヘテロポリマー, heteropolymer)もある。")
118
分子病(molecular disease) :
ヘモグロビン異常症は遺伝子の突然変異によりヘモグロビン分子の構造異常が起こった状態であるが このような遺伝性のタンパク質分子構造異常による病気を分子病と呼ぶ。

119
分子レベルで明らかにされた最初の分子病である。
鎌状赤血球貧血症 N-val · his · leu · thr · pro · glu · glu · · · · ·C(146) HbS β鎖 HbA β鎖 N-val · his · leu · thr · pro · val · glu · · · · ·C(146) 分子レベルで明らかにされた最初の分子病である。
 HbS β鎖. HbA β鎖. N-val · his · leu · thr · pro · val · glu · · · · ·C(146) 分子レベルで明らかにされた最初の分子病である。")
120
たった1個の極性残基が非極性残基に置換されたのことによってヘモグロビン分子が凝集し、これが繰り返してゲル状の大きな重合体となり、さらに重合して不溶性の結晶となるため赤血球の鎌状化が起こる。鎌状の赤血球が破壊されやすいので貧血を引き起こす。

121
五、タンパク質の分類 * タンパク質の組成による分類 単純タンパク質 複合タンパク質 アポタンパク質 + 補欠分子族
* タンパク質の組成による分類 単純タンパク質 複合タンパク質 アポタンパク質 + 補欠分子族 * タンパク質全体の形による分類 繊維状タンパク質 球状タンパク質

122
分類 補欠分子族 例を挙げて 核タンパク質 核 酸 染色体 糖タンパク質 糖 質 免疫グロブリン リポタンパク質 脂 質 β‒リポタンパク質 リンタンパク質 リン酸 カゼイン 色素タンパク質 色 素 ヘモグロビン 金属タンパク質 金属イオン フェリチン

123
The Relation of Structure and Function of Protein
第 三 節 タンパク質構造と機能の関係 The Relation of Structure and Function of Protein

124
一、タンパク質一次構造と機能の関係 (一)一次構造が空間構造の基礎 ジ ス ルフィ ド 結合 ウシ膵臓リボヌクレアーゼ
一次構造が空間構造の基礎 ジ ス ルフィ ド 結合 ウシ膵臓リボヌクレアーゼ")
125
天然状態, 触媒活性あり 尿素、β-MEを除去する 尿素、 β-ME 変性,活性なく

126
二、タンパク質空間構造と機能の関係 (一)ミオグロビンとヘモグロビンの構造 目 录
ミオグロビンとヘモグロビンの構造 目 录")
127
(三)タンパク質コンホメーション 変化と病気 タンパク質コンホメーション変化による 病気: タンパク質のホールデイグ(folding)が誤った場合ではその一次構造が変化しないにもかかわらずコンホメーション変化が起こり タンパク質機能に影響を及ぼす。
が誤った場合ではその一次構造が変化しないにもかかわらずコンホメーション変化が起こり タンパク質機能に影響を及ぼす。 .")
128
狂牛病の原因は異常プリオン(prion protein, PrP)と呼ばれる感染力のあるタンパク質である。
狂牛病(mad cow disease)は人及び動物がともに罹患する神経退行性病気である。 狂牛病の原因は異常プリオン(prion protein, PrP)と呼ばれる感染力のあるタンパク質である。
は人及び動物がともに罹患する神経退行性病気である。 狂牛病の原因は異常プリオン(prion protein, PrP)と呼ばれる感染力のあるタンパク質である。")
129
正常なPrPcがα-螺旋に富み,病気に罹った場合β‒プリ‒ツ•シ‒トに変換する(PrPsc )
狂牛病

130
第 四 節 タンパク質の物理的化学的性質 と分離精製
The Physical and Chemical Characters and Separation and Purification of Protein

131
一、物理的化学的性質 (一)タンパク質の両性解離
タンパク質は実際には生理的pHでは荷電されており、それはC末端とN末端以外に極性のある側鎖によるものである。

132
例えば Lysのε‒NH2、Argのグアニジノ基、Hisのイミダゾール基などはプロトンを受け入れて正電荷(陽イオン)を持っており,AspとGluの非α‒COOHはプロトンを失って負の電荷(陰イオン)をもっている。
を持っており,AspとGluの非α‒COOHはプロトンを失って負の電荷(陰イオン)をもっている。")
133
*タンパク質の等電点(isoelectric point, pI)
陽イオン 両性イオン 陰イオン pH< pI pH=pI pH>pI *タンパク質の等電点(isoelectric point, pI) あるpHにおいてはタンパク質分子の正味の電荷が0となり、直流電場に置かれても移動しない。この溶液のpH値をタンパク質の等電点と呼ぶ。
 あるpHにおいてはタンパク質分子の正味の電荷が0となり、直流電場に置かれても移動しない。この溶液のpH値をタンパク質の等電点と呼ぶ。")
134
直流電場において荷電粒子はそれらの符号と異にする極へ向かって移動する。このような現象を電気泳動(electrophoresis)と呼ぶ。
と呼ぶ。")
135
生体内におけるタンパク質の大部分のpIは弱酸性に偏っているので電気泳動を行う場合ではアルカリ性条件がよく用いられる。イオン交換クロマトグラフィー場合では陰イオン交換剤がよく用いられる。
(二)タンパク質のコロイド性質
タンパク質のコロイド性質.")
136
*タンパク質コロイドを安定させる因子 • 顆粒表面に帯びた電荷 • 水和層
タンパク質分子の大きさはコロイド粒子の範囲(1-100nm)に属する。タンパク質溶液をコロイド溶液と見なしている。球状タンパク質は水によく溶けて親水性コロイドである。 *タンパク質コロイドを安定させる因子 • 顆粒表面に帯びた電荷 • 水和層
に属する。タンパク質溶液をコロイド溶液と見なしている。球状タンパク質は水によく溶けて親水性コロイドである。 *タンパク質コロイドを安定させる因子. • 顆粒表面に帯びた電荷. • 水和層.")
137
溶液中のタンパク質の沈殿 水和層 - + + - 正電荷を帯びた タンパク質 等電点にある タンパク質 負電荷を帯びた タンパク質 アルカリ
正電荷を帯びた タンパク質 等電点にある タンパク質 - 負電荷を帯びた タンパク質 アルカリ 酸 酸 アルカリ 脱水作用 + 正電荷を帯びた タンパク質 - 負電荷を帯びた 安定しないタンパク質 酸 アルカリ 溶液中のタンパク質の沈殿

138
(三)タンパク質の変性、沈殿と凝析 * タンパク質の変性(denaturation) 定義 タンパク質分子は種々の原因により一次構造が変化せずに高次構造のみ とくに水素結合が破壊され その物理的、化学的性質が変化し、生物活性が喪失される。
タンパク質の変性、沈殿と凝析 * タンパク質の変性(denaturation) 定義 タンパク質分子は種々の原因により一次構造が変化せずに高次構造のみ とくに水素結合が破壊され その物理的、化学的性質が変化し、生物活性が喪失される。")
139
化学的:極端な酸性、アルカリ、重金属塩、アルカロイド試薬、有機溶媒,尿素 などの変性剤,界面活性化剤など
変性の実質 タンパク質の天然コンホメーションの破壊 変性の原因 物理的:加熱、放射線、超音波,撹はん 化学的:極端な酸性、アルカリ、重金属塩、アルカロイド試薬、有機溶媒,尿素 などの変性剤,界面活性化剤など

140
変性による変化: • 溶解度の減少 • 粘度の増加 • 生物活性の喪失 • タンパク質分解酵素の作用受けやすさ • 結晶能力の喪失

141
*応用 臨床医学では 変性因子がよく消毒及び滅菌に用いられる。 その外に 変性を防ぐのはタンパク質製品(例えば ワクチンなど)を保存する必要条件である。 。
を保存する必要条件である。 。")
142
タンパク質の再生(renaturation)
ある程度で変性したタンパク質は変性剤を除去した後もとの高次構造、生物活性などを回復することができる。 タンパク質の再生を確かめたのは1957年Christean Anfinsenのウシ膵臓リボヌクレアーゼ(Rnase)での見事な実験である。
での見事な実験である。")
143
天然状態, 触媒活性あり 尿素、β-MEを除去する 尿素、 β-ME 変性,活性なく

144
* タンパク質の沈殿 よく 用いられる沈殿試薬: • 中性塩類 • 有機溶媒 • アルカロイド • 重金属(Ag,Pb,Hg)などがある。 塩析(salt precipitation) タンパク質溶液に多量の中性塩類をくわえて溶解度を減少させて沈殿を生ずる現象である。

145
* タンパク質の凝析(protein coagulation)
タンパク質は強酸、強アルカリの作用で変性を起こす。その時強酸、強アルカリの溶液の中に溶けている。もし溶液のpH をタンパク質の等電点に調和させば変性タンパク質が不溶性のものとなる。さらに加熱によって硬い凝塊となり,もはや 強酸、強アルカリの溶液の中に溶けていない。

146
280nmに最大吸収を持つ。これは芳香族アミノ酸であるTrp,Tyr残基に共役二重結合があることに起因する。
(四)タンパク質の紫外線吸収 紫外線吸収スペクトルの特徴: 280nmに最大吸収を持つ。これは芳香族アミノ酸であるTrp,Tyr残基に共役二重結合があることに起因する。 したがって280nmでの吸光度(A280,OD280)の測定はタンパク質の定量によく用いられる。
タンパク質の紫外線吸収. 紫外線吸収スペクトルの特徴: 280nmに最大吸収を持つ。これは芳香族アミノ酸であるTrp,Tyr残基に共役二重結合があることに起因する。 したがって280nmでの吸光度(A280,OD280)の測定はタンパク質の定量によく用いられる。")
147
(五)タンパク質の呈色反応 ⒈ ニンヒドリン反応(ninhydrin reaction) タンパク質の分解によって生じたアミノ酸はニンヒドリンと反応する。 ⒉ ビウレット反応(biuret reaction) アルカリ性の条件の下でタンパク質におけるペプチド結合は銅イオン(Cu2+)と錯体を形成し 赤紫色を呈する。
と錯体を形成し 赤紫色を呈する。")
Similar presentations









 ~基本的なことについて~.>")






>")
 分子生物学概観>")
 脂質 たんぱく質 プラス五大栄養素 ビタミン 無機質.>")


