細胞と多様性の 生物学 第3回 転写と翻訳 和田 勝 東京医科歯科大学教養部
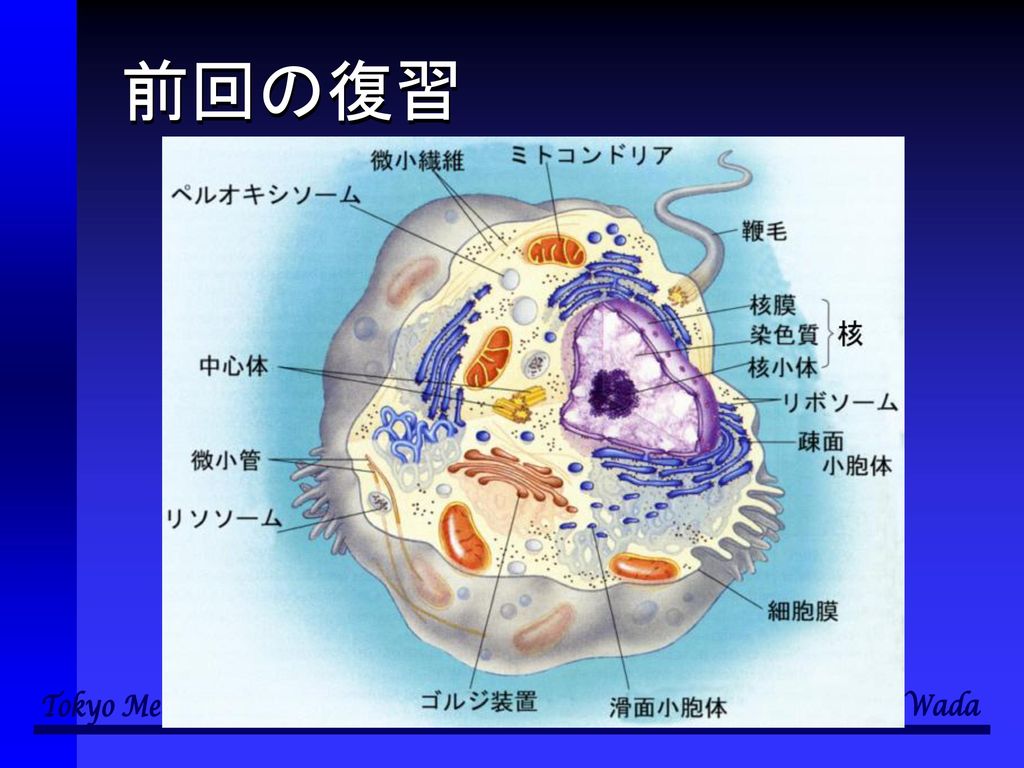
前回の復習
核(nucleus) ●核膜 ●核小体 ●染色質
染色体-染色質-DNA ●ヌクレオソーム ●染色体 ●凝縮した染色質 ●DNA 遺伝情報 ●染色質
染色体-染色質-DNA 染色体 染色質 (chromatin) DNA 細胞分裂のとき 遺伝情報の読み出しのとき
遺伝情報の流れ 設計図(遺伝情報、DNA) 遺伝情報とタンパク質の仲立ちとなるRNA タンパク質が構造と機能を実現 転写(transcription) 遺伝情報とタンパク質の仲立ちとなるRNA 翻訳(translation) タンパク質が構造と機能を実現 (セントラル・ドグマ)
生体高分子 細胞を構成する分子は、巨大分子(macromolecule)である。 いずれも単位となるモノマーの重合体(ポリマー)である。 いずれも単位となるモノマーの重合体(ポリマー)である。 DNA タンパク質 多糖類 ヌクレオチド アミノ酸 単糖 (グルコース、ガラクトースなど)
DNAが遺伝情報 DNAはポリヌクレオチドである。 ヌクレオチド=ヌクレオシド+リン酸
ヌクレオシド ヌクレオシド=デオキシリボース+塩基 (五炭糖) 5’ 1’ 4’ 3’ 2’
塩基 アデニン(A) プリン塩基 グアニン(G) シトシン(C) ピリミジン塩基 チミン (T) (ウラシル(U))
ヌクレオチドの伸長 5’ 伸長は 5’→ 3’方向に起こる フォスフォジエステル結合 3’
DNAの一本鎖 糖の部分から塩基が突き出ている形となる。 DNAの片方の鎖骨格部分 (5’)P-S-P-S-P-S-P-S-P-S-P-S(3’) (Pはリン酸、Sは糖) 糖の部分から塩基が突き出ている形となる。
DNAの一本鎖 塩基は4種で、シトシン(C)、チミン(T)アデニン(A)、グアニン(G)。 塩基は4種で、シトシン(C)、チミン(T)アデニン(A)、グアニン(G)。 糖とリン酸の骨格部分はどの部分でも共通、違いは塩基の配列の順序。 省略すると、DNAの片方の鎖は CTAGTCGTA....... のように、5’側から3’に向かって書くことができる。
DNAは二本鎖 二本鎖の向かい合った2つの塩基の間に水素結合ができる =O: H-N- このような2つの基が近づくと =O---H-N-
DNAは二本鎖 AとT (2本) GとC (3本)
DNAは二重ラセン
DNA分子
DNAの性質 二本の鎖は逆平行(anti-parallel) AとT、GとCが、それぞれ二本と三本の水素結合で結合 多数の水素結合のため強い結合となるが、共有結合ではないので必要なときには少ないエネルギーで切れる 片方の鎖の塩基配列が決まれば、もう片方は一意に決まる
DNAからRNAへ 設計図(遺伝情報、DNA) 遺伝情報とタンパク質の仲立ちとなるRNA タンパク質が構造と機能を実現 転写(transcription) 遺伝情報とタンパク質の仲立ちとなるRNA 翻訳(translation) タンパク質が構造と機能を実現 (セントラル・ドグマ)
転写の過程 5’→3’に書かれた遺伝情報は、これに相補的な鎖を鋳型にして、RNAへ転写される
DNAからRNAへ コード鎖、センス鎖、有効鎖、sense strandと言う 5' ATGGAATTCTCGCTC…….. 3' 3' TACCTTAAGAGCGAG……. 5' DNA二本鎖 読み取り方向 鋳型鎖、アンチセンス鎖、antisense strandと言う 転写されたmRNA 5' AUGGAAUUCUCGCUC 3'
mRNA このRNA転写物を伝令RNA (messenger RNA, mRNA)という。 合成した短いRNA(たとえばUUUUUUUUU..….)がどんなポリペプチドを作るか調べた。
mRNA→アミノ酸の配列 UUUUUUUUU..….はPhe-Phe-Phe-….だった。 3つの塩基が1つのアミノ酸を指定することが確かめられた。 3つの塩基(コドンという)の64の組み合わせに対応するアミノ酸の暗号表が明らかになった。
Second Position of Codon A G Fi r s t Po s i t i o n TTT Phe [F] TTC TTA Leu [L] TTG TCT Ser [S] TCC TCA TCG TAT Tyr [Y] TAC TAA Ter [end] TAG TGT Cys [C] TGC TGA TGG Trp [W] Th i r d Po s i t i o n CTT CTC CTA CTG CCT Pro [P] CCC CCA CCG CAT His [H] CAC CAA Gln [Q] CAG CGT Arg [R] CGC CGA CGG ATT Ile [I] ATC ATA ATG Met [M] ACT Thr [T] ACC ACA ACG AAT Asn [N] AAC AAA Lys [K] AAG AGT AGC AGA AGG GTT Val [V] GTC GTA GTG GCT Ala [A] GCC GCA GCG GAT Asp [D] GAC GAA Glu [E] GAG GGT Gly [G] GGC GGA GGG
DNAは文字列である DNAはATCG(あるいはコドン)という文字でアミノ酸の並び方を記録した文字列のようなものである。 必要に応じて本を開いて、必要な部分をmRNAに書き写して、核の外へ送り出す。
DNAは文字列である DNA分子としてはATCGが切れ目なく続くので、読み初めと、終わりを明示する必要がある。 読み初めは、Metを指定するコドンと同じATG。終わり(終止コドン)はTAA, TAG, TGAである。
DNAの複製 体細胞分裂の過程で必ずDNAの複製(DNA replication)がおこる。 DNAの塩基は、AとT、GとCが相補的に結合するので、片方のDNA鎖に対しては一意にもう片方のDNA鎖が決まってしまう。
DNAの複製 このような複製方法は、「半保存的な」複製と呼ばれている。
DNAの複製の過程 複製開始点で二本鎖がほどける 「複製装置」が形成される DNAポリメラーゼがヌクレオチドを伸長していく
DNAの5’→3’伸長
リーディング鎖とラギング鎖
校正と修復 複製の過程で誤りの起こる確立は非常に低く抑えられているが、これはDNAポリメラーゼに校正機能が備わっているからで、誤りがあるとそれを正している。 さまざまの誤りや変異を修復する機構が備わっている(DNA修復、DNA repair)。
核小体 核小体では、rRNA遺伝子の転写ががおこり、細胞質でつくられたタンパク質といっしょになり、次に登場するリボソームが作られている。
途中のまとめ すべて細胞には核があり、核の中にはDNAがある。DNAはコドンという文字で、タンパク質の設計図を書いた本である。
タンパク質の合成 設計図(遺伝情報、DNA) 遺伝情報とタンパク質の仲立ちとなるRNA タンパク質が構造と機能を実現 転写(transcription) 遺伝情報とタンパク質の仲立ちとなるRNA 翻訳(translation) タンパク質が構造と機能を実現 (セントラル・ドグマ)
お話の舞台
RNA RNA(ribonucleic acid)とDNAの違いは 1)糖がデオキシリボースではなく リボースであること(五員環の2’ リボースであること(五員環の2’ に水酸基がつく) 2)塩基としてチミンのかわりにウ ラシルが使われること 3)二本鎖ではなく一本鎖であること
RNA RNAには3種類ある 1)mRNA(messenger RNA) DNAの情報をコピーしてリボソーム へもってくる へもってくる 2)rRNA(ribosomal RNA) リボソームの構成要素 3)tRNA(transfer RNA) 特定のアミノ酸を合成の場へ運搬
tRNAの構造 特定のアミノ酸を3’に結合 一本鎖内に水素結合 アミノ酸に対応したアンチコドン
リボソームの構造
リボソームの構造
翻訳の過程
翻訳の過程
翻訳の開始 メチオニンを結合した開始tRNAが開始因子とともにリボソーム小単位と結合する。 ついでこれらがmRNAと5’末端に結合し、3’に沿って読み始めのAUGを探す。 開始tRNAはP部位に結合し、開始因子が離れ、大単位が結合する。
翻訳の終了 リボソームのA部位に終止コドンが来ると、終結因子が結合する。 アミノ酸の代わりに水が付加され、完成したペプチド鎖はリボソームから離脱する。 リボソームとmRNAは解離する。
塩基の配列→アミノ酸の配列 読み初めのMetを指定するコドン(AUG)から、終止コドン(UAA, UAG, UGA)まで、順番にアミノ酸をつないでいく。 塩基の配列がアミノ酸の配列に翻訳される。
アミノ酸 R NH2-C-COOH H H R O H-N+-C-C H H O- アミノ酸の構造式 アミノ酸の構造式 (中性水溶液中)
アミノ酸の種類 Asp D Ala A Glu E Gly G Arg R Val V Lys K Leu L His H Ile I アスパラギン酸 Asp D 負電荷 アラニン Ala A 非極性 グルタミン酸 Glu E グリシン Gly G アルギニン Arg R 正電荷 バリン Val V リシン Lys K ロイシン Leu L ヒスチジン His H イソロイシン Ile I アスパラギン Asn N 極性 プロリン Pro P グルタミン Gln Q フェニルアラニン Phe F セリン Ser S メチオニン Met M トレオニン Thr T トリプトファン Trp W チロシン Tyr Y システイン Cys C
ペプチド結合 H R1 O H-N+-C-C H H O- H R2 O H-N+-C-C H H O- + H2O H R1 O R2 O H-N+-C- C-N- C-C H H H O- ペプチド結合
ポリペプチド アミノ末端(N末端) カルボキシル末端(C末端)
タンパク質の一次構造 アミノ酸の配列が決まると、タンパク質の性質はおのずと決まる。 アミノ酸の配列のことを、タンパク質の一次構造(primary structure)という。 アミノ末端側を左にして順番に書き、番号を付ける。
タンパク質の一次構造 たとえば卵白リゾチームの一次構造は次のとおりである。 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 1 Lys Val Phe Gly Arg Cys Glu Leu Ala Ala Ala Met Lys Arg His 15 16 Gly Leu Asp Asn Tyr Arg Gly Tyr Ser Leu Gly Asn Trp Val Cys 30 31 Ala Ala Lys Phe Glu Ser Asn Phe Asn Thr Gln Ala Thr Asn Arg 45 46 Asn Thr Asp Gly Ser Thr Asp Tyr Gly Ile Leu Gln Ile Asn Ser 60 61 Arg Trp Trp Cys Asn Asp Gly Arg Thr Pro Gly Ser Arg Asn Leu 75 76 Cys Asn Ile Pro Cys Ser Ala Leu Leu Ser Ser Asp Ile Thr Ala 90 91 Ser Val Asn Cys Ala Lys Lys Ile Val Ser Asp Gly Asn Gly Met 105 106 Asn Ala Trp Val Ala Trp Arg Asn Arg Cys Lys Gly Thr Asp Val 120 121 Gln Ala Trp Ile Arg Gly Cys Arg Leu
タンパク質の二次構造 二次構造(secondary structure)には、2種類ある αヘリックス(α-helix) βシート(β-sheet) 逆平行βシート 平行βシート
αへリックス Ball and stick模型
αへリックス リボン模型
βシート Ball and stick模型
βシート リボン模型
チャイム Chimeを使った図を見せます。Chimeは下記のサイトからダウンロードして、インストールすることができます(無料)。 http://www.mdlchime.com/chime/ あるいは講談社ブルーバックス「パソコンで見る動く分子事典」B1266の付録CD-ROMから同上のプラグインをインストールしてください。 Chimeはブラウザーのプラグインで、分子の構造式を表示し、マウスによって回転して、いろいろな角度から分子を見たり、さまざまな表現方法を選んだりと、いろいろな使い道があります。
タンパク質のドメイン αヘリックスやβシートなどの二次構造が組み合わせって、球状となった構造単位があることがわかってきた。 大きなタンパク質を構築するモジュール単位で、これをドメインと呼ぶ。
タンパク質の三次構造 サイトゾール中では、側鎖の性質のために決まった立体構造(conformation)をとる。
タンパク質の四次構造 一本のポリペプチド鎖を一つの単位として、これが複数組み合わさったタンパク質がある。これを四次構造という。 たとえばヘモグロビンはα鎖2本とβ鎖2本の四量体である。 α2β2
二つの合成経路
分泌性タンパク質の合成 先頭にシグナルペプチドが付加している。膜タンパク質の場合も同じ。
分泌タンパク質の一次構造 翻訳直後の、卵白リゾチームの一次構造は次のとおり 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 1 Met Arg Ser Leu Leu Ile Leu Val Leu Cys Phe Leu Pro Leu Ala 15 16 Ala Leu Gly Lys Val Phe Gly Arg Cys Glu Leu Ala Ala Ala Met 30 31 Lys Arg His Gly Leu Asp Asn Tyr Arg Gly Tyr Ser Leu Gly Asn 45 46 Trp Val Cys Ala Ala Lys Phe Glu Ser Asn Phe Asn Thr Gln Ala 60 61 Thr Asn Arg Asn Thr Asp Gly Ser Thr Asp Tyr Gly Ile Leu Gln 75 76 Ile Asn Ser Arg Trp Trp Cys Asn Asp Gly Arg Thr Pro Gly Ser 90 91 Arg Asn Leu Cys Asn Ile Pro Cys Ser Ala Leu Leu Ser Ser Asp 105 106 Ile Thr Ala Ser Val Asn Cys Ala Lys Lys Ile Val Ser Asp Gly 120 121 Asn Gly Met Asn Ala Trp Val Ala Trp Arg Asn Arg Cys Lys Gly 135 136 Thr Asp Val Gln Ala Trp Ile Arg Gly Cys Arg Leu オレンジ色がシグナルペプチド
まとめと次回 今回はタンパク質の生合成と構造を理解し、機能分子であるタンパク質について学んだ。 今回はタンパク質の生合成と構造を理解し、機能分子であるタンパク質について学んだ。 次回は、細胞が生きていくために必要なエネルギーをどのように得ているかなどを見ていこう。