Download presentation
1
生 体 酸 化 Biological Oxidation
第 六 章 生 体 酸 化 Biological Oxidation

2
*生体酸化の概念 生体内で行われた物質の酸化反応を生体酸化(biological oxidation)と呼ぶ。 主に糖質,脂肪,タンパク質などは生体内で酸化作用を通じて最後CO2, H2Oとエネルギ-を生成する 過程を指す。 糖質 脂肪 タンパク質 O2 CO2とH2O ADP+Pi エネルギ- ATP 熱

3
*生体酸化と体外燃焼の同じところ 生体酸化の様式には脱電子,脱水素及び酸素添加反応がある。そのうち 脱電子、脱水素反応は 最もよく 見られる。酸化還元反応の一般規則に遵っている。 消費された酸素量も最終産物(CO2,H2O)も放出されたエネルギーも みな同じである。
も放出されたエネルギーも みな同じである。")
4
*生体酸化と体外燃焼の違い 生 体 酸 化 体 外 燃 焼 環境条件 pHがほぼ中性に近く,37℃,水がある環境、
生 体 酸 化 体 外 燃 焼 環境条件 pHがほぼ中性に近く,37℃,水がある環境、 強酸、強アルカリ,高温,乾燥 反応様式 すべて酵素で触媒する。 しだいに進行する。 O2が電子を受け取ってから水素イオンと化合してH2Oを生成する。CO2は脱炭素反応によって生成される。 触媒が必要ではない激烈、爆発的に進行H2とCは直接O2と化合してH2OとCO2を生成する。 エネルギ-放出 段階的に放出する。その一部分が化学エネルギ-の形即ちATPに捕捉されて貯えられる。 突然,一次的に放出される。全部 熱や光の形で放出される。

5
*生体酸化の一般過程 アセチルCoA TAC Gn TG タンパク質 グルコース FA+グリセロール アミノ酸 CO2 ADP+Pi ATP
2H H2O 呼吸鎖

6
第一節 ATPを生成する酸化系 The Oxidation System of ATP Producing

7
一、呼吸鎖 定義 一定の順序で配列されている水素伝達体と電子伝達体は細胞の酸素を取り入れるという呼吸過程に密接に関係しているので呼吸鎖(respiratory chain)と呼ぶ。呼吸鎖は電子伝達鎖 (electron transfer chain)ともいう。
と呼ぶ。呼吸鎖は電子伝達鎖 (electron transfer chain)ともいう。 .")
8
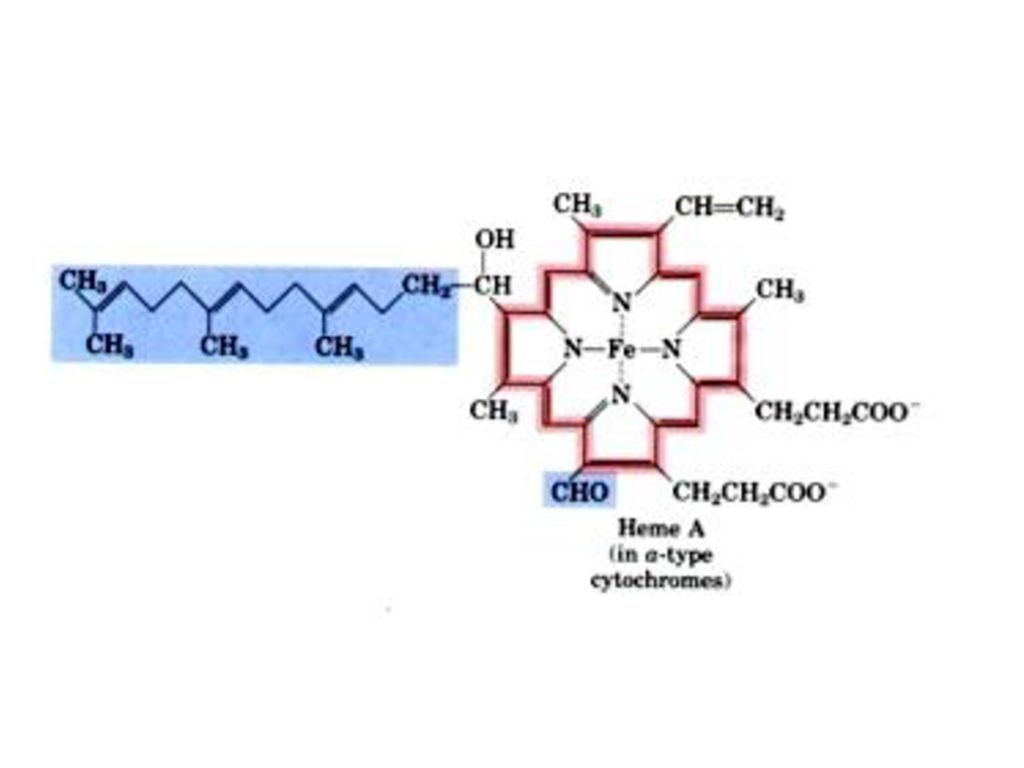
* CoQとCyt c は上述した呼吸鎖複合体に含まれない。
(一)呼吸鎖の組成 四つの電子伝達体複合体 (complex) ヒトのミトコンドリアにある呼吸鎖複合体 複合体 酵 素 補欠分子族 複合体Ⅰ NADH-CoQ還元酵素 (NADH脱水素酵素) FMN, Fe-S 複合体Ⅱ コハク酸-CoQ還元酵素 (コハク酸脱水素酵素) FAD, Fe-S 複合体Ⅲ CoQ-Cyt C還元酵素 鉄-ポルフィリン、 Fe-S 複合体Ⅳ Cyt Cオキシダーゼ 鉄-ポルフィリン, Cu * CoQとCyt c は上述した呼吸鎖複合体に含まれない。
呼吸鎖の組成. 四つの電子伝達体複合体 (complex) ヒトのミトコンドリアにある呼吸鎖複合体. 複合体. 酵 素 補欠分子族. 複合体Ⅰ. NADH-CoQ還元酵素. (NADH脱水素酵素) FMN, Fe-S. 複合体Ⅱ. コハク酸-CoQ還元酵素. (コハク酸脱水素酵素) FAD, Fe-S. 複合体Ⅲ. CoQ-Cyt C還元酵素. 鉄-ポルフィリン、 Fe-S. 複合体Ⅳ. Cyt Cオキシダーゼ. 鉄-ポルフィリン, Cu. * CoQとCyt c は上述した呼吸鎖複合体に含まれない。")
10
ミトコンドリアの構造

11
ミトコンドリア内膜にある呼吸鎖複合体の位置

12
Q e- Cytc 細胞質ゾル側 Ⅲ 糸粒体内膜 Ⅱ Ⅳ Ⅰ フマル酸 コハク酸 マトリック側 NADH+H+ NAD+ 1/2O2+2H+
H2O

13
1.複合体Ⅰ : NADH‐CoQ 還元酵素 働き: 電子をNADHからCoQ (ubiquinone,ユビキ ノン)に転移する 複合体Ⅰ
FMN; Fe-SN-1a,b; Fe-SN-4; Fe-SN-3; Fe-SN-2

14
NAD+(CoⅠ)とNADP+(CoⅡ)は多くの脱水酵素の補酵素として役割を果たす。両者はVit PPの誘導体と見なす。
NAD+,NADP+では酸化還元反応に関与する部分はピリジン環である。ピリジン環の1位窒素は五価で電子を受けると三価窒素に転換できる。またH原子がピリジン環の4位のCに可逆的に結合することができる。したがって酸化還元反応においてNAD+或はNADP+は水素運搬体として働く。

15
NAD+とNADP+の構造 R=H: NAD+; R=H2PO3:NADP+

16
NAD+(NADP+)とNADH(NADPH)の相互変換 NAD+とNADP+は1個の水素原子と1電子を受け取る。
とNADH(NADPH)の相互変換 NAD+とNADP+は1個の水素原子と1電子を受け取る。")
17
NAD+或はNADP+を補酵素とする脱水素酵素
酵 素 補 酵 素 イソクエン酸脱水素酵素 NAD+ or NADP+ α-ケトグルタ-ル脱水素酵素 NAD+ グルセルアルデヒド-3-リン酸 脱水素酵素 リンゴ酸脱水素酵素 乳酸脱水素酵素 グルコ-ス-6-リン酸脱水素酵素 NADP+ グルタミン酸脱水素酵素 NAD+or NADP+

18
NADH或はNADPHは340nmの所に特徴的光吸収をもつので酵素反応の測定に広く用いられる。

19
FMN: VitB2即ちリボフラビンの誘導体である。酸化還元反応に関与する部分はリボフラビンのイソアロキサジン環である。

20
リボフラビンを補欠分子族とする酵素 酵 素 補欠分子族 アシル-CoA脱水素酵素 FAD コハク酸脱水素酵素 ジヒドロリポイル 脱水素酵素 グリセロ-ル-3-リン酸 NADH脱水素酵素 FMN

21
鉄-硫黄タンパク質: 非ヘム鉄と酸不安定性硫黄を含む。 Fe2S2と Fe4S4としてよく見られる。 単電子伝達体

22
鉄-硫黄タンパク質 S 無機硫黄 Cys-硫黄

23
ユビキノン(ubiquinone, coenzyme Q, CoQ):
セミキノン型 (遊離基) 全酸化型または キノン型 還元型または キノール型 (ヒドロキノン)
 全酸化型または. キノン型. 還元型または. キノール型. (ヒドロキノン)")
24
還元型または キノール型 (ヒドロキノン) セミキノン型 (遊離基) 全酸化型または キノン型
 セミキノン型 (遊離基) 全酸化型または キノン型")
25
NADH+H+ NAD+ FMN FMNH2 還元型Fe-S 酸化型Fe-S Q QH2 複合体Ⅰの機能 マトリックス側 細胞質ゾル側

26
Structure of NADH-Q Oxidoreductase (Complex I)
Structure of NADH-Q Oxidoreductase (Complex I). The structure, determined by electron microscopy at 22-Å resolution, consists of a membrane-spanning part and a long arm that extends into the matrix. NADH is oxidized in the arm, and the electrons are transferred to reduce Q in the membrane.
. The structure, determined by electron microscopy at 22-Å resolution, consists of a membrane-spanning part and a long arm that extends into the matrix. NADH is oxidized in the arm, and the electrons are transferred to reduce Q in the membrane.")
27
2.複合体Ⅱ:コハク酸-CoQ還元酵素 働き :電子をコハク酸からCoQに転移する 复合体Ⅱ コハク酸→ →CoQ
Fe-S1; b560; FAD; Fe-S2 ; Fe-S3

28
シトクロム類(cytochromes, Cyt)
一群ヘム含有タンパク質である。呼吸鎖組成に 関与するCytは3種類ある。単電子伝達体である。

30
細胞質ゾル側 マトリックス側 コハク酸

31
3.複合体Ⅲ: CoQ- Cyt c還元酵素 働き:電子をCoQからCyt cに渡す。 複合体Ⅲ QH2→ →Cyt c
b562; b566; Fe-S; c1

32
細胞質ゾル側 マトリックス側

33
4.複合体Ⅳ : Cyt cオキシダーゼ 働き:電子をCyt c からO2に渡す。
CuA→a→a3→CuB そのうちCyt a3とCuBからなっている活性部位は電子をO2に渡す。

34
細胞質ゾル側 マトリックス側

35
(二)呼吸鎖成分の配列順序 次の実験デ-タによって定められたのである。 ①標準酸化還元電位の測定 ②呼吸鎖の分解及び組換え
③特異的阻害剤による吸収スペクトルの変化 ④体外のミトコンドリア実験

36
呼吸鎖における各酸化還元対の標準酸化還元電位
E0’(V)
")
37
1. NADH酸化呼吸鎖 2.コハク酸酸化呼吸鎖 NADH → 複合体Ⅰ →Q →複合体Ⅲ→Cyt c →複合体Ⅳ→O2

38
NADH酸化呼吸鎖 FADH2酸化呼吸鎖 コハク酸

39
電子伝達鎖 コハク酸 複合体Ⅱ 複合体Ⅰ 複合体Ⅲ 複合体Ⅳ

40
二、酸化的リン酸化 *定義 基質からの還元当量は電子伝達系によってO2をH2Oに還元させる。ATPシンタ-ゼはこの過程で放出されたエネルギ-を利用してADPとPiからATPを合成する。このような酸化反応と密接に共役するリン酸化を酸化的リン酸化(oxidative phosphorylation) と呼ぶ。
 と呼ぶ。")
41
基質に含まれるエネルギ-を直接にADP(GDP)に転移し,ATP(GTP)を生成する様式を基質レベルのリン酸化(substrate level phosphorylation)と呼ぶ。
に転移し,ATP(GTP)を生成する様式を基質レベルのリン酸化(substrate level phosphorylation)と呼ぶ。")
42
(一)酸化的リン酸化の共役部位 酸化的リン酸化の共役部位 : 複合体Ⅰ、Ⅲ、Ⅳ P/O 比と自由エネルギ-変化に基づいて
複合体Ⅰ、Ⅲ、Ⅳ P/O 比と自由エネルギ-変化に基づいて ⊿Gº'=-nF⊿Eº' P/O 比: 1グラム原子酸素が消費された時に消費された無機リンのグラム原子の値を指す。つまり 合成されたATPのモル数を示す。

43
によって測定されたいくつかの基質のP/O比
体外のミトコンドリア実験 によって測定されたいくつかの基質のP/O比 基 質 呼 吸 鎖 の 組 成 P/O比 ATP生成数 β‐ヒドロキシ酪酸 NADH→FMN→CoQ→ Cyt→O2 2.4~2.8 3 コハク酸 FAD→CoQ→Cyt→O2 1.7 2 アスコルビン酸 Cyt c→Cyt aa3→O2 0.88 1 Cyt C Cyt aa3→O2 0.61~0.68

44
電子伝達鎖における自由エネルギ-変化 コハク酸 酸化的リン酸化の共役部位 部位 ATPが産生できるか否か 産生できる 産生できる 産生できる
電位変化 自由エネルギー変化 部位 ATPが産生できるか否か 産生できる 産生できる 産生できる コハク酸 酸化的リン酸化の共役部位 ATP ATP ATP

45
(二) 酸化的リン酸化の共役機構 酸化的リン酸化の共役を説明するのに2種類の仮説 即ち化学仮説と化学浸透圧仮説がある。化学仮説はどのリン酸化部位においても酸化とリン酸化が直接化学に共役していると仮定する。酸化とリン酸化をつないでいるエネルギ-の高い中間体を仮定するのである。しかし この中間体はこれまで実際に分離されたことがなく,そのため この仮説はいまではあまり信用されていない。

46
1、化学浸透圧仮説 Mitchellの化学浸透圧仮説 (chemiosmotic hypothesis) は呼吸鎖で起こる酸化とリン酸化が共役するメカニズムを以下のように説明する。 呼吸鎖成分の酸化によって生じた水素イオン(プロトン、H+)がまずミトコンドリア内膜の外側に放出され、これによって膜内外のプロトン濃度に差ができる。続いて,この濃度差に基づく電気化学的ポテンシャルを利用してATPが合成される。
がまずミトコンドリア内膜の外側に放出され、これによって膜内外のプロトン濃度に差ができる。続いて,この濃度差に基づく電気化学的ポテンシャルを利用してATPが合成される。")
47
化学浸透圧仮説 マトリックス ミトコンドリア膜 ADP + Pi ATP O2 H2O H+ e- H+

48
化学浸透圧仮説 細胞質ゾル側 ミトコンドリア内膜 マトリックス側

51
化学浸透圧仮説 - - - - - - - - - 細胞質ゾル側 + + + + + + + + + + Q マトリックス側 H+ Ⅲ Ⅰ
Ⅱ Ⅳ F0 F1 Cyt c Q H+ フマル酸 コハク酸 NADH+H+ NAD+ 1/2O2+2H+ H2O マトリックス側 ADP+Pi ATP

52
The path of electrons through Complex III probably involves a "Q cycle“
such as that shown here (blue arrows).
.")
53
Q Cycle:

55
Path of electrons through Complex IV

56
呼吸鎖複合体Ⅰ、Ⅲ、Ⅳはみなプロトンポンプとして働いている。

57
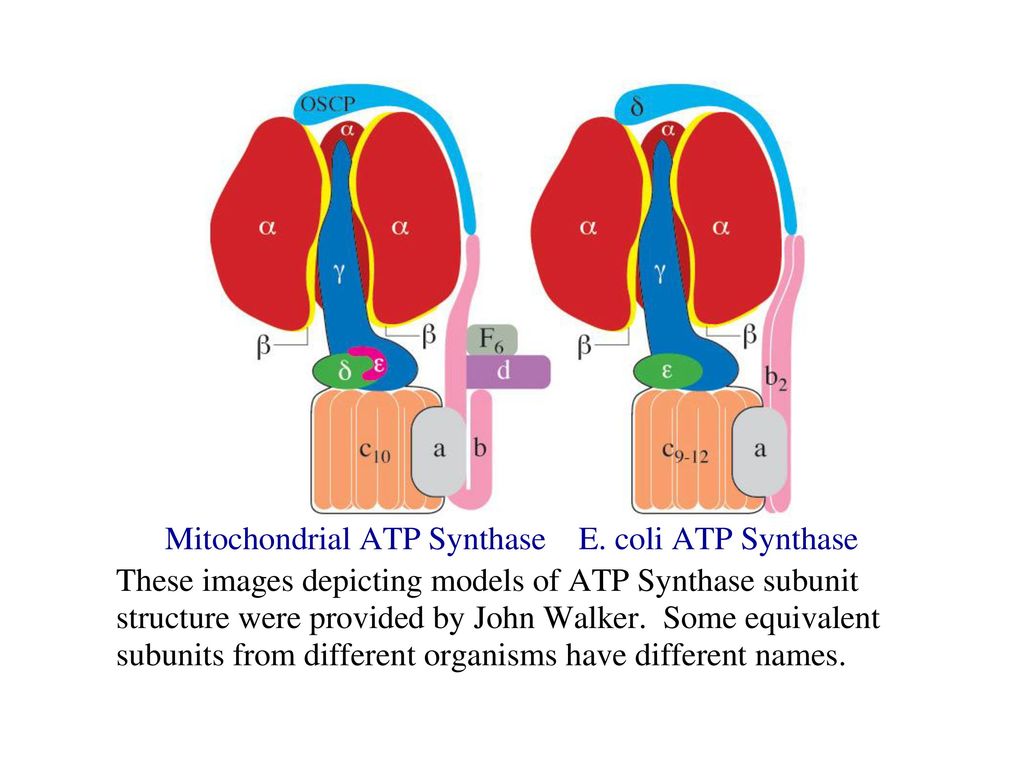
2. ATPシンタ-ゼ (ATP synthase)
ATPシンタ-ゼは呼吸鎖複合体Ⅴと見なされており,主に 二つの機能単位であるF0とF1からなっている。 ATPシンタ-ゼの構造

58
F1は親水性であり,膜の内側に突き出し, 5種類のサブユニット 即ちα,β,γ,δとεからなる(α3β3γδε)。βが触媒活性をもつがαと結合してからはじめて触媒活性を示す。
ATPシンタ-ゼの構造

59
F0は疎水性であり,内膜に埋められ,a1b2c9~12サブユニットからなり,プロトンチャンネルの一部分を構成している。
ATPシンタ-ゼの構造

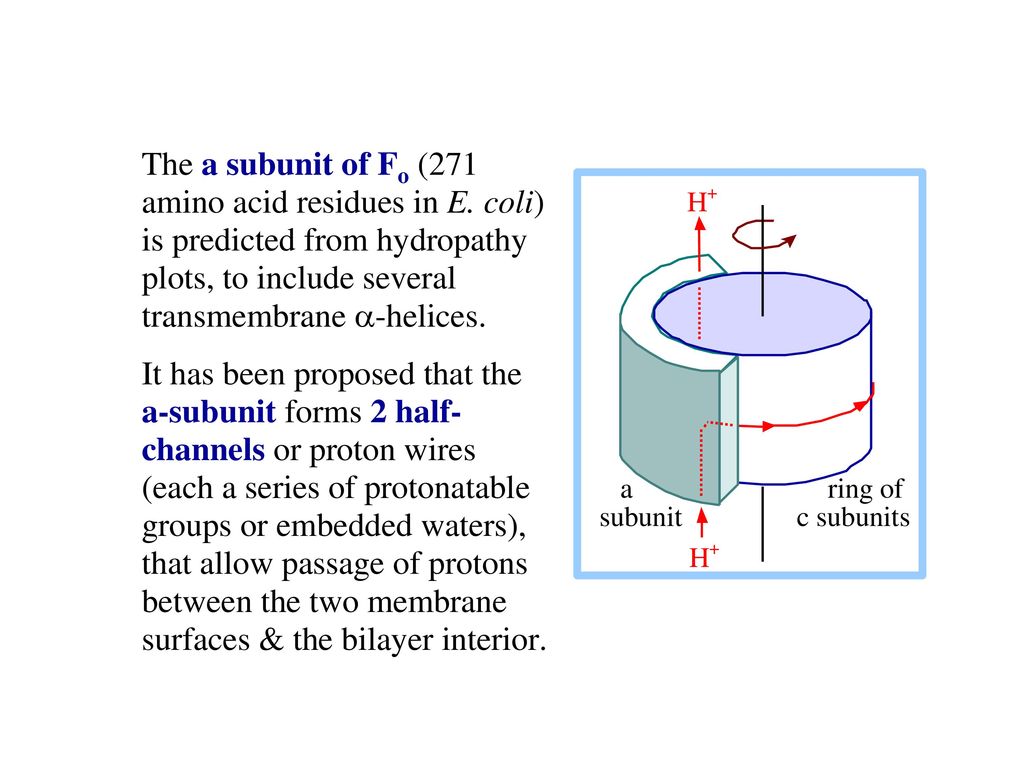
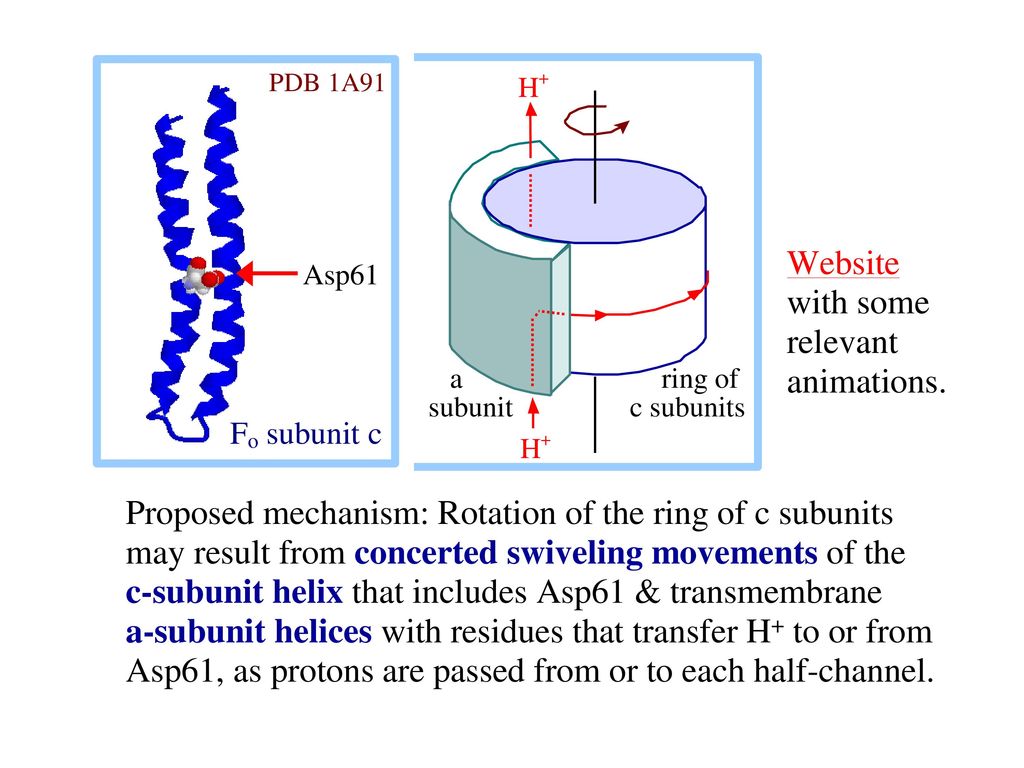
64
H+が濃度勾配に従ってF0のaとcの間を通っている時,γが回旋し, 三つのβサブユニットの立体配置が変化する。
ATPシンタ-ゼの働きの機構

66
F1Fo couples ATP synthesis to H+ transport into the mitochondrial matrix. Transport of least 3 H+ per ATP is required.

67
三、酸化的リン酸化に影響を及ぼす因子 (一)阻害剤 1. 呼吸鎖阻害剤 呼吸鎖のある特定部位で電子伝達を阻害する。
呼吸鎖のある特定部位で電子伝達を阻害する。 2. 脱共役剤(uncoupler) リン酸化反応を阻害するが電子伝達を阻害しない 例えば:2,4-ジニトロフェノ-ル(2,4-DNP) 3.酸化的リン酸化阻害剤 電子伝達とリン酸化を両方とも阻害する。 例えば:オリゴマイシン
 リン酸化反応を阻害するが電子伝達を阻害しない. 例えば:2,4-ジニトロフェノ-ル(2,4-DNP) 3.酸化的リン酸化阻害剤. 電子伝達とリン酸化を両方とも阻害する。 例えば:オリゴマイシン.")
68
呼吸鎖阻害剤による阻害部位 コハク酸 CO,CN-, アンチマイシンA N3-とH2S ジメルカプロ-ル × × × ロテノン
アモバルビタ-ル

69
異なった基質と阻害剤がミトコンドリアにおける酸素消費に影響する。
ピ ル ビ ン 酸 異なった基質と阻害剤がミトコンドリアにおける酸素消費に影響する。 ADP(過量) コ ハ ク 酸 ミ ト コ ン ド リ ア DNP ロ テ ノ ン オリ ゴマ イシ ン アンチマイシン 時間
 コ. ハ. ク. 酸. ミ. ト. コ. ン. ド. リ. ア. DNP. ロ. テ. ノ. ン. オリ. ゴマ. イシ. ン. アンチマイシン. 時間.")
70
脱共役タンパク質の作用機構(褐色脂肪組織) 熱 H+
細胞質 ゾル側 Ⅳ Cyt c UCP Ⅰ Ⅲ F0 F1 Q Ⅱ マトリック ス側 ADP+Pi ATP

71
F0に作用してH+の内部流入をブロックする。
オリゴマイシン(oligomycin) F0に作用してH+の内部流入をブロックする。 細胞質ゾル側 マトリックス側 オリゴマイシン ATPシンターゼ構造
 F0に作用してH+の内部流入をブロックする。 細胞質ゾル側. マトリックス側. オリゴマイシン. ATPシンターゼ構造.")
72
(二)ADPの調節作用 (三)甲状腺ホルモンの作用 (四)ミトコンドリアDNAの突然変異
呼吸制御率(respiratory control ratio, RCR) (三)甲状腺ホルモンの作用 Na+,K+–ATPア-ゼと脱共役タンパク質遺伝子の発現がいずれも増加する。 (四)ミトコンドリアDNAの突然変異 ミトコンドリアDNA病気及び衰老に関連する。
 (三)甲状腺ホルモンの作用. Na+,K+–ATPア-ゼと脱共役タンパク質遺伝子の発現がいずれも増加する。 (四)ミトコンドリアDNAの突然変異. ミトコンドリアDNA病気及び衰老に関連する。")
73
コハク酸 電子伝達鎖と酸化的リン酸化の概貌 ΔμH+ プロトン電気化学勾配,H+m:マトリックス側,H+c: 細胞質ゾル側

74
四、ATP 高リン酸エネルギ-結合と高エネルギ-リン酸化合物 高エネルギ-リン酸結合
加水分解された時そのΔG0’が-21KJ/molより負であるリン酸エステル結合をいう。Pで表す。 高エネルギ-リン酸化合物 高エネルギ-リン酸結合をもっている。

75
よく見られる高エネルギ-化合物 -43.9 -61.9 -41.8 -30.5 -31.4 共通式 例を挙げ 放出されたエネルギ-
(pH7.0,25℃)KJ/mol ホスホクレアチン (ホスホグアニジン) -43.9 ホスホエノ-ル ピルビン酸(PEP) (エノ-ルリン酸) -61.9 アセチルリン酸 -41.8 ATP,GTP,UTP,CTP -30.5 アセチルCoA -31.4
KJ/mol. ホスホクレアチン. (ホスホグアニジン) ホスホエノ-ル. ピルビン酸(PEP) (エノ-ルリン酸) アセチルリン酸 ATP,GTP,UTP,CTP アセチルCoA")
76
ヌクレオシド二リン酸キナーゼの作用 ATP + UDP ADP + UTP ATP + CDP ADP + CTP ATP + GDP ADP + GTP アデニル酸キナーゼの作用 ADP + ADP ATP + AMP

77
CP:筋肉と脳組織におけるエネルギーの貯蔵形の一つである。
クレアチンキナ-ゼの作用 クレアチンキナ-ゼ クレアチン ホスホクレアチン(CP) CP:筋肉と脳組織におけるエネルギーの貯蔵形の一つである。
 CP:筋肉と脳組織におけるエネルギーの貯蔵形の一つである。")
78
ATPの生成と利用 ATP ADP ~P 酸化的リン酸化 ~P ATPは細胞のエネルギ-交換の中で中心的な役割を果たしている。 クレアチン
ホスホク レアチン 機械的仕事(筋肉収縮) 濃度勾配に逆らう物質輸送 (能動輸送) 化学的エネルギ-(生合成反応) 電気エネルギ-(生物電気) 熱エネルギ-(体温の維持) ADP 酸化的リン酸化 基質レベルのリン酸化 ATPは細胞のエネルギ-交換の中で中心的な役割を果たしている。
 濃度勾配に逆らう物質輸送. (能動輸送) 化学的エネルギ-(生合成反応) 電気エネルギ-(生物電気) 熱エネルギ-(体温の維持) ADP. 酸化的リン酸化. 基質レベルのリン酸化. ATPは細胞のエネルギ-交換の中で中心的な役割を果たしている。")
79
五、ミトコンドリア内膜を通る 物質の輸送 ほとんどの親水性物質はミトコンドリア内膜を通過できないので特異的な輸送系が必要である。。多くの物質はミトコンドリア内膜における交換輸送体(transporter,トランスポーター)によって輸送される。
によって輸送される。")
80
ミトコンドリア内膜における主な輸送体系

81
(一)細胞質ゾル中のNADHの酸化 ミトコンドリアの外部で生じたNADHが呼吸鎖によって酸化されるには ミトコンドリア内膜を透過しなければならない。ミトコンドリア内膜にはNADH輸送系がないので 還元当量をシャトルという巧妙な仕掛けでミトコンドリア内に送り込む。
細胞質ゾル中のNADHの酸化 ミトコンドリアの外部で生じたNADHが呼吸鎖によって酸化されるには ミトコンドリア内膜を透過しなければならない。ミトコンドリア内膜にはNADH輸送系がないので 還元当量をシャトルという巧妙な仕掛けでミトコンドリア内に送り込む。")
82
1. グリセロ-ル-3-リン酸シャトル 主に脳と骨格筋にある。

83
NADH+H+ DHAP 呼吸鎖 グリセロ-ル -3-リン酸 脱水素酵素 FADH2 FAD グリセロ-ル -3-リン酸 NAD+ 糸粒体
外膜 糸粒体 内膜 マトリ ックス 膜間腔

84
2.リンゴ酸―アスパラギン酸シャトル これは主に肝臓と心臓にある。

85
呼吸链 NADH +H+ NAD+ NADH +H+ 糸粒体内膜 NAD+ 細胞質ゾル マトリックス Glu-Asp 輸送体 Asp OAA
GOT Glu リンゴ酸脱 水素酵素 NADH +H+ NAD+ NADH +H+ 糸粒体内膜 α-KG NAD+ リンゴ酸 リンゴ酸-α-KG 輸送体 細胞質ゾル マトリックス

86
(二)アデニル酸運搬体 (adenine nucleotide transporter)
アデニル酸運搬体 (adenine nucleotide transporter)")
87
H+ ADP3- ADP3- ATP4- H+ H2PO4- H2PO4- ATP4-
F0 F1 細胞質ゾル側 マトリッ クス側 アデニル 酸輸送体 リン酸 輸送体 ATP4-

88
F1Fo couples ATP synthesis to H+ transport into the mitochondrial matrix. Transport of least 3 H+ per ATP is required.

89
Phosphate reenters the matrix with H+ by an electro-neutral symport mechanism. Pi entry is driven by, & uses up, the pH gradient (equivalent to one mol H+ per mol ATP).
..")
90
第二節 その他の酸化系 The Others Oxidation Enzyme Systems
第二節 その他の酸化系 The Others Oxidation Enzyme Systems

91
一、好気性脱水素酵素オキシダーゼ 水素受容体 補助因子 産物 嫌気性脱水素酵素 補酵素 好気性脱水素酵素 O2 FMN FAD H2O2
Cu H2O

92
二、ペルオキシソームにおける酸化酵素類 (一)カタラーゼ(catalase) 補欠分子族は4個のヘムを含む。 カタラーゼ 2H2O + O2
カタラーゼ(catalase) 補欠分子族は4個のヘムを含む。 カタラーゼ 2H2O + O2")
93
(二)ペルオキシダーゼ(perioxidase)
ヘム補欠分子族をとし, H2O2をフェノール或はアミン類に酸化される。 R + H2O2 RO + H2O RH2+ H2O2 R + 2H2O ペルオキシダーゼ

94
三、スーパーオキシドジスムターゼ (superoxide dismutase)
反応性酸素分子 スーパーオキシド陰イオン遊離基(O2﹣), H2O2,ヒドロキシフリーラジカル(•OH)の総称 SOD 2O2﹣+ 2H+ H2O2 + O2 カタラーゼ H2O + O2
, H2O2,ヒドロキシフリーラジカル(•OH)の総称. SOD. 2O2﹣+ 2H+ H2O2 + O2. カタラーゼ. H2O + O2.")
95
3.セレンを含むグルタチオンペルオキシダーゼ
H2O2 (ROOH) 2G –SH NADP+ グルタチオン 還元酵素 グルタチオンペルオキシダーゼ H2O (ROH+H2O) NADPH+H+ G –S – S – G * 生体膜及びヘモグロビンはこの酵素によってその損傷を免れる。
 2G –SH. NADP+ グルタチオン. 還元酵素. グルタチオンペルオキシダーゼ. H2O. (ROH+H2O) NADPH+H+ G –S – S – G. * 生体膜及びヘモグロビンはこの酵素によってその損傷を免れる。")
96
(一)モノオキシゲナーゼ(monoxygenase)
四、ミクロソームにおける酸化酵素類 (一)モノオキシゲナーゼ(monoxygenase) *触媒された反応: RH + NADPH + H+ + O2 ROH + NADP+ + H2O 混合機能酸化酵素(mixed-function oxidase)或は水酸化酵素(hydroxylase,ヒドロキシラーゼ)ともいう。 反応においては Cyt P450を必要とする。
モノオキシゲナーゼ(monoxygenase) *触媒された反応: RH + NADPH + H+ + O2. ROH + NADP+ + H2O. 混合機能酸化酵素(mixed-function oxidase)或は水酸化酵素(hydroxylase,ヒドロキシラーゼ)ともいう。 反応においては Cyt P450を必要とする。")
97
水酸化酵素による反応

98
(二)ジオキシゲナーゼ 2個酸素原子を二重結合の二つの炭素原子に添加させる。 例えば (O2) Trpピロラーゼ
ジオキシゲナーゼ 2個酸素原子を二重結合の二つの炭素原子に添加させる。 例えば (O2) Trpピロラーゼ")









 炭水化物の消化・吸収 食環境デザインコース 3年 09210211~09210220.>")



-イ-aーH.>")


 ~基本的なことについて~.>")





n -COOH R-(CH2)n-CO-S-CoA R-(CH2)n-CO-S-CoA CoA-SH>")
解糖系はほとんどすべての生物に共通に存在する糖の代謝経路である。 2)反応は細胞質で行われる。>")
 脂質 たんぱく質 プラス五大栄養素 ビタミン 無機質.>")