Download presentation
Presentation is loading. Please wait.

1
分子コンピューティング ─計算機科学者(素人)から見た ナノテクノロジーと 分子エレクトロニクス─
萩谷 昌己 東京大学・大学院情報理工学系研究科 JST CREST

2
Special Thanks 有賀先生(ERATO相田ナノ空間) 岩崎先生・吉信先生(阪大) 西川先生(大阪電通大) 陶山先生(東大)
特定領域「分子プログラミング」の皆様 JST CREST 「多相的分子インタラクション」の皆様

3
あらすじ ボトムアップ・ナノテクノロジーと DNAナノテクノロジー(その現状は?) 分子コンピューティングと
分子プログラミング(その目標は?) 真の分子メモリと 分子アドレシング(その心は?) 真の分子メモリと分子アドレシングはCRESTのプロジェクト。 ここで、最初の話題と繋がる。 (なぞかけ研究者)
 真の分子メモリと. 分子アドレシング(その心は?) 真の分子メモリと分子アドレシングはCRESTのプロジェクト。 ここで、最初の話題と繋がる。 (なぞかけ研究者)")
4
ボトムアップ・ナノテクノロジーと DNAナノテクノロジー (その現状は?)
トップダウン 従来の加工技術や計測技術をスケールダウン 各種リソグラフィ SPM(Scanning Probe Microscopy) ボトムアップ 自己組織化
 ボトムアップ. 自己組織化.")
5
情報処理 2002 Vol.43 No.1 特集 ナノテクのトレンド
量子電子デバイスの最近のトピックス 日立ケンブリッジ研究所 中里 和郎 超分子化学からのナノテクノロジーへの アプローチ 科学技術振興事業団 相田ナノ空間プロジェクト 有賀 克彦 トップダウン? ボトムアップ

6
トップダウンとボトムアップ トップダウン ボトムアップ パターンを微細化していく方法 リソグラフィとエッチング SPM
フォト・電子ビーム・イオンビーム SPM ボトムアップ 原子や分子をナノ構造に集積させる方法 SPM? 自己組織化 分子線エピタキシー 蒸着法 分子線エピタキシーや蒸着法は、自己組織化のための方法ではないか。 (ナノテクノロジー・ ハンドブック 日経BP)
")
7
SPMはボトムアップ? SPM(Scanning Probe Microscopy) 原理的にはトップダウン
STM(Scanning Tunneling Microscopy) AFM(Atomic Force Microscopy) 原理的にはトップダウン しかし、リソグラフィとは違って、 個々の分子を直接的に操作している。 大量生産できない。 cf. 多探針STM
 AFM(Atomic Force Microscopy) 原理的にはトップダウン. しかし、リソグラフィとは違って、 個々の分子を直接的に操作している。 大量生産できない。 cf. 多探針STM.")
8
http://alme1.almaden.ibm.com/vis/stm/atomo.html Iron on Copper
Carbon Monoxide on Platinum Xenon on Nickel

9
AFM陽極酸化による微細パターン作成 (阪大・産研・岩崎研究室)
Si + 2H2O SiO2 + 4H+ + 4e–

10
AFM酸化システム

11
AFM酸化システム

12
AFM酸化膜パターンの形成例

13
タンパクのポジティブ・パターンの作成法

14
ポジティブ・パターンの作成例 5mm 5mm 1mm 1mm

15
AFMによる量子ドットの作成 (富士通研究所) http://pr.fujitsu.com/jp/news/2002/07/29.html
AFMによる局所酸化と分子線エキタピキー選択成長技術の組み合わせ。 量子ドットの最小直径は20nm。

16
なぜボトムアップか? トップダウンは限界なのか?
進歩するトップダウン技術 遠紫外リソグラフィ 電子ビーム:EB(Electron Beam) イオンビーム:FIB(Focused Ion Beam) 生き伸びるMooreの法則 ボトムアップと組み合わせるにしても、 トップダウンの技術は必要。
 イオンビーム:FIB(Focused Ion Beam) 生き伸びるMooreの法則. ボトムアップと組み合わせるにしても、 トップダウンの技術は必要。")
17
インテル・リソグラフィ・ロードマップ The Intel Lithography Roadmap, May 16, 2002
DUVはDeep Ultra-Violetで、波長は248nm EUVは、13nm。 加工線幅は、1999年で130nm。ちなみに、集積密度は、2000年で400万トランジスタ。 2009年には、extreme ultra-violet lithography、遠紫外リソグラフィにより、32nmになる。 The Intel Lithography Roadmap, May 16, 2002

18
遠紫外リソグラフィ EUV露光装置のプロトタイプ。 The Intel Lithography Roadmap, May 16, 2002

19
Moore predicts more advances
BBC News Tuesday, 11 February, 2003, 10:12 GMT Moore predicts more advances "It will be a real challenge," Dr Moore told the 50th anniversary meeting of the International Solid-State Circuits Conference in San Francisco. "No exponential is forever. Your job is to delay forever." チップに集積されるトランジスタの数は2年おきに2倍になる。 ムーアの法則はあと10年は続くだろう。しかしそのためには多大な努力が必要になる。 世界最大の半導体メーカーIntel社の創設者の一人である Gordon Moore博士が1965年に経験則として提唱した、 「半導体の集積密度は18~24ヶ月で倍増する」という法則。 この法則によれば、半導体の性能は指数関数的に向上していくことになる。 実際には、集積密度の向上ペースはこれより鈍化しているが、 「集積密度」を「性能向上」に置き換えて考えると、この法則は現在でも成立しているとされ、 今後の半導体の性能向上を予測する際の指標として広く用いられている。 このことから、この法則を「半導体の性能向上は…」と表現する例もある。 ただし、ムーアの法則は半導体の微細加工技術の発展を根拠としているため、 2010年代には微細化が原子レベルにまで到達してしまい、ムーアの法則は通用しなくなると予想されている。

20
それでも分子エレクトロニクス? やはりトップダウンの限界 単電子デバイス(量子ドット)の自然な実現 これまでに、分子素子の様々な提案
delay foreverのために(特にMooreの第二法則) 単電子デバイス(量子ドット)の自然な実現 超省電力の単電子トランジスタなど? 将来的に、量子計算のための素子??? これまでに、分子素子の様々な提案 例えば、HP QSR(Quantum Science Research) しかし、どうやって配線するのか、という大問題 しかし、専門家に言わせれば、 派生技術は実り多いが、 ブレークスルーが必要(確実な見通しは無い)。 自己組織化!
 単電子デバイス(量子ドット)の自然な実現. 超省電力の単電子トランジスタなど? 将来的に、量子計算のための素子??? これまでに、分子素子の様々な提案. 例えば、HP QSR(Quantum Science Research) しかし、どうやって配線するのか、という大問題. しかし、専門家に言わせれば、 派生技術は実り多いが、 ブレークスルーが必要(確実な見通しは無い)。 自己組織化!")
21
ロタクサン C. P. Collier et al. Science 285, 391-394 (1999)
この他、単一分子素子として有名なのは、 1974年にIBMのAviramとRatnerが提案した分子ダイオード 1988年にAviramが提案した分子トランジスタ 三つの繋がったベンゼン環の真中がねじれるやつ カーボンナノチューブ C60分子(フラーレン)を使った単電子トランジスタ(Wada他、2000) C. P. Collier et al. Science 285, (1999)
を使った単電子トランジスタ(Wada他、2000) C. P. Collier et al. Science 285, (1999)")
22
カテナン C. P. Collier et al. Science 289, 1172-1175 (2000) +2Vでopen
-2Vでclose ~0.1Vでread UCLA ⇒ HP C. P. Collier et al. Science 289, (2000)
")
23
自己組織化というけれど… 人によって千差万別。 粒子やドット、もしくは、その配列の形成を 意味することが多い。 自己組織化の場も色々。
液相・気相・界面・膜・・・ 回路のような複雑なパターンを 自己組織化させる試みは決して多くはない。 DNAナノテクノロジー プログラムされた自己組織化 Algorithmic self-assembly

24
量子ドットの自己組織化 (電通大・山口浩一研究室) http://crystal.ee.uec.ac.jp/dot.html
GaAs上にInAsの微小な結晶粒を自己組織化。 SK成長モード(Stanski-Krastnaow)と呼ばれる成長条件(歪の利用) 高さ6nm、底辺20nm。 透過型電子顕微鏡
と呼ばれる成長条件(歪の利用) 高さ6nm、底辺20nm。 透過型電子顕微鏡.")
25
タンパクによる量子ドット配列の自己組織化 (松下電器・山下一郎研究室+奈良先端大・冬木研究室)
タンパクによる量子ドット配列の自己組織化 (松下電器・山下一郎研究室+奈良先端大・冬木研究室) 直径12nmのフェリチンに直径6nmの粒子を貯蔵させる。
 直径12nmのフェリチンに直径6nmの粒子を貯蔵させる。")
26
分子の鎖でナノワイヤー配線 (理研・青野研究室)
ジアセチレン化合物分子 ワイアの太さは3nm

27
ここでちょっと脇道 量子ドット・コンピュータ
キワモノか? 量子コンピュータとは違う。 量子ドット・セル・オートマトン(QCA) ドミノのように四個の 量子ドットを並べる。 ドミノ内ではトンネル効果に よって電子が移動。 ドミノの相互作用により 状態が伝搬する。 配線が必要無い? しかし、量子ドットを正確に 配置しなければならない。 QCAによるインバータ ドミノはセルという。 セルには四個の量子ドットと二個の電子。 隣り合うセル内の電子はクーロン力によって相互作用する。
 ドミノのように四個の. 量子ドットを並べる。 ドミノ内ではトンネル効果に. よって電子が移動。 ドミノの相互作用により. 状態が伝搬する。 配線が必要無い? しかし、量子ドットを正確に. 配置しなければならない。 QCAによるインバータ. ドミノはセルという。 セルには四個の量子ドットと二個の電子。 隣り合うセル内の電子はクーロン力によって相互作用する。")
28
DNAナノテクノロジー DNAによる自己組織化
DNAの網 分子糊としてのDNA DNAによるナノ粒子の自己組織化 DNAによるナノワイアの自己組織化 DNAタイル DNA自身による構造形成 プログラムされた自己組織化

29
ここで、DNAの大きさについて Seeman先生 二本鎖の直径は2nm。 一回転が10~10.5ベースペア。
スタックの幅が0.34nm、従って、一回転が3.5nmくらい。 Seeman先生

30
DNAの網 (阪大・産研・川合研究室) SiO2/Si基盤 親水性のあるSiO2基板上にのみネットワークを形成 写真はμメートル四方

31
DNAによるナノ粒子の自己組織化初期の研究
C. A. Mirkin et al. DNA-based method for rationally assembling nanoparticles into macroscopic materials. Nature 382, (1996) A. P. Alivisatos et al. Organization of ‘nanocrystal molecules’ using DNA. Nature 382, (1996)
 A. P. Alivisatos et al. Organization of ‘nanocrystal molecules’ using DNA. Nature 382, (1996)")
32
ナノ粒子:C. A. Mirkin et al. Nature (1996)
直径13nm

33
ナノ粒子:A. P. Alivisatos et al. Nature (1996)
")
34
ナノ粒子からワイヤへ E. Braun et al. Nature (1998)
電極間の距離は12~16μm。 λ-DNA ワイヤの長さは12μm、直径は100nm。 銀イオン

35
ナノワイアの自己組織化 N. I. Kovtyukhova and T. E. Mallouk
Nanowires as Building Blocks for Self-Assembling Logic and Memory Circuits Chem. Eur. J., 8, (2002) ワイアの直径は15nmから350nm。
 ワイアの直径は15nmから350nm。")
36
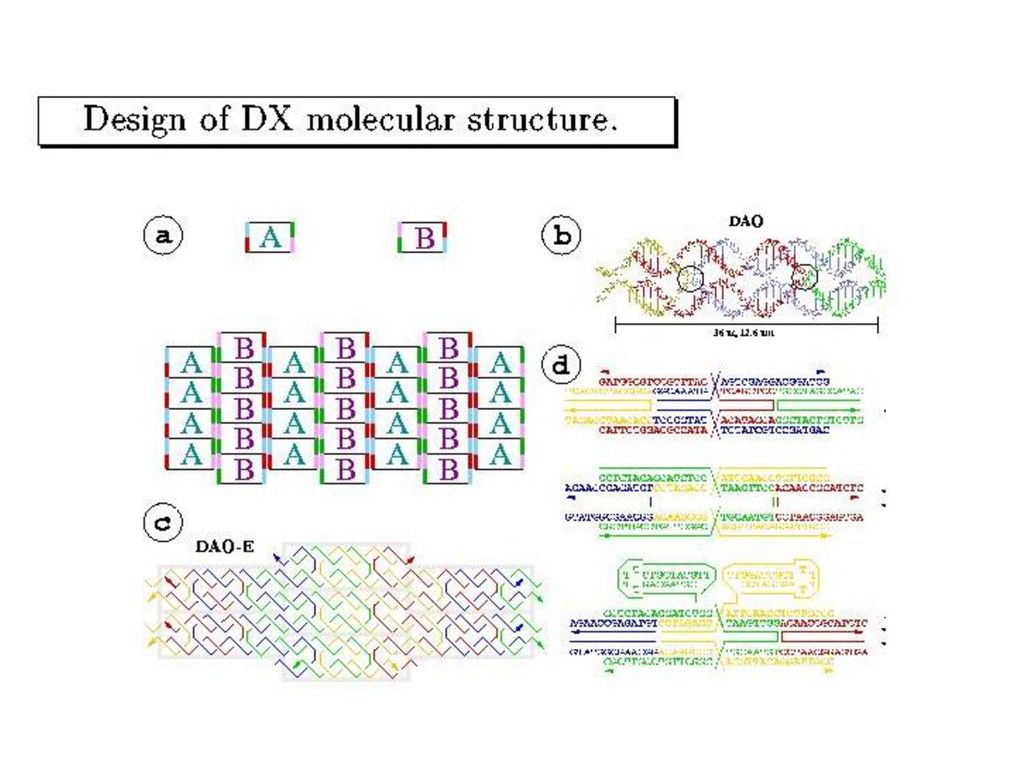
Winfree-SeemanのDNAタイル
(double crossover分子)
")
38
棒は300nm。

39
The experimental result will be presented at DNA9, 2003.

40
Journal of Nanoparticle Research 4: 313-317 (2002)
金のナノパーティクル 直径は1.4nm S. Xiao et al. Journal of Nanoparticle Research 4: (2002)
")
41
Nadrian Seeman New York University, Professor (chemistry)
1995 Feynman Prize in Nanotechnology

42
妄想 自己組織化された分子回路の ビンディング・ブロック(たとえばadderとか) いかにして既存の半導体回路に組み込むか 自己組織化の制御
巧妙にエネルギーを注入することにより、 状態を遷移させて非可逆的に自己組織化を進める。 分子マシン研究の動機の一つ 「精製」のプロセス エラーを含む分子の除去 いかにして既存の半導体回路に組み込むか ボトムアップとトップダウンのハイブリッド

43
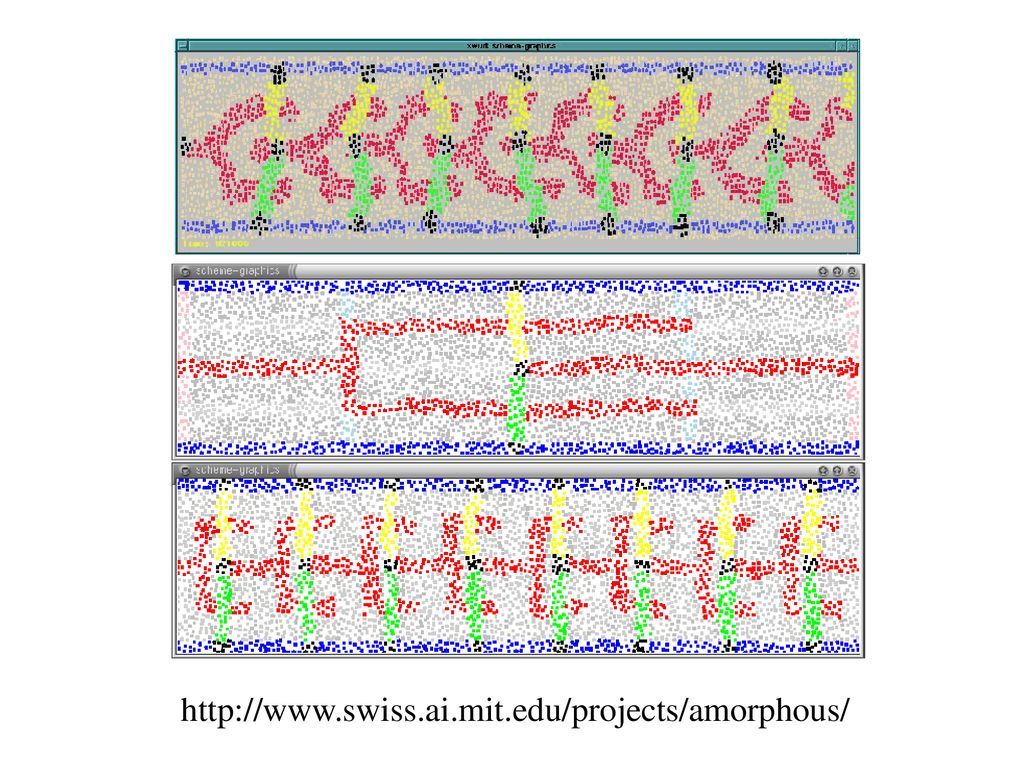
また脇道:アモルファス計算 自己組織化のための新しい計算パラダイム Computational particle
微細加工技術と細胞工学 低コストで様々なプロセッサ Computational particle 小さい計算力と少量メモリー 不規則配置、可動性 非同期、局地的相互作用 誤った挙動、環境の影響 同一プログラム 自分たちの位置や方向に 関する情報をもたない 近接のparticleと短距離 (半径r)の通信をする。 全体としては超並列計算システムになっている。 回路の自己組織化のシミュレーション
の通信をする。 全体としては超並列計算システムになっている。 回路の自己組織化のシミュレーション.")
45
ボトムアップとトップダウンの ハイブリッド
自己組織化された分子回路を いかにして既存の半導体回路に組み込むか カーボンナノチューブ DNA 量子ドットによるフローティング・ゲート型メモリ インプリント・リソグラフィによるクロスワイア 分子回路に見合うトップダウン技術 例えば、FIBやインプリント・リソグラフィ

46
カーボンナノチューブを用いた 量子効果ナノデバイスの集積化技術 (産総研+富士通研究所)
電極幅は20μ。 電極間は4μ。 鉄をパターニング。 メタンガスを900度、30分。

47
導線としてのDNA http://chem.ch.huji.ac.il/~porath/ 電極間は8nm。
10nm、30bpのDNAの電導率を計測。

48
フローティング・ゲート型メモリ (松下電器・山下一郎研究室+奈良先端大・冬木研究室)
直径12nmのフェリチンに直径6nmの粒子を貯蔵させる。

49
クロスワイア(HP) http://www.hpl.hp.com/news/2002/oct-dec/beyond_silicon.html
クロスワイアはμ四方。 電子ビームとインプリント。

50
HP

51
必要とされる人材 (ここまでのまとめに代えて)
計算機科学、特に量子計算を含む 各種計算理論に精通していて、 半導体物理の深い知識を持つと同時に、コンピュータ・シミュレーションを駆使して 分子素子の設計を行い、 しかも実際に有機合成して、 SPMによる観測までもやってしまう人。 そんな奴、いるわけないだろ! 極めて学際的な分野。 スーパーマンが必要。

52
分子コンピューティングと 分子プログラミング (その目標は?)
生体分子=情報処理装置 化学反応の自律的制御 → 自身にコード化 (組み合わせ的複雑さ) 分子コンピューティング 潜在的計算能力の理学的解明 新しい計算機能の工学的実現 分子プログラミング 分子計算のシステマティックな設計論 新しい情報処理パラダイムの構築 生命科学・微小科学への計算概念の移転
 分子コンピューティング. 潜在的計算能力の理学的解明. 新しい計算機能の工学的実現. 分子プログラミング. 分子計算のシステマティックな設計論. 新しい情報処理パラダイムの構築. 生命科学・微小科学への計算概念の移転.")
53
設計論のための3つの切り口 進化 高信頼性 (設計方法論) (計算過程) 自己組織化 (計算原理) タンパク工学 ドラッグデザイン
バイオテクノロジー 医療応用 ナノテクノロジー 分子エレクトロニクス タンパク工学 ドラッグデザイン 自己組織化 (計算原理) 要するに、分子コンピューティングの応用。
 要するに、分子コンピューティングの応用。")
54
分子コンピューティング 様々な計算モデル(とその解析) Adleman-Lipton Seeman-Winfree
DNAの選択的ハイブリダイゼーションを利用した 解の候補のランダムな生成 データ並列計算による解の抽出 Suyama --- dynamic programming Sakamoto-Hagiya --- SAT Engine Seeman-Winfree 各種形態のDNA分子の自己組織化(self-assembly) 自己組織化による計算
 自己組織化による計算.")
55
Adlemanの最初のDNAコンピュータ
O0 O6 PCR PAGE AF-SEP O1 O2 O3 O4 O5 HP

56
Adleman-Liptonパラダイム 全ての解候補の生成 解の検査と抽出 解の検出 全ての 代入の生成 (Lipton 1995,
充足可能性問題) 0 0 0 0 変数Xn 変数X1 変数X2 変数X3 1 1 1 1 解の検査と抽出 Ti : 文字列の多重集合 (試験管) 解の検出 [Detect 命令] [Separate命令] T2 =+(T1, s) : s を含む配列の抽出 T2 =-(T1, s) : s を含まない配列の抽出 [Merge命令] T3 =T1 U T2 : T1 と T2 の混合 [Amplify命令] (T2, T3) =T1 : T1 の増幅(コピー)
 0. 0. 0. 0. 変数Xn. 変数X1. 変数X2. 変数X3. 1. 1. 1. 1. 解の検査と抽出. Ti : 文字列の多重集合 (試験管) 解の検出. [Detect 命令] [Separate命令] T2 =+(T1, s) : s を含む配列の抽出. T2 =-(T1, s) : s を含まない配列の抽出. [Merge命令] T3 =T1 U T2 : T1 と T2 の混合. [Amplify命令] (T2, T3) =T1 : T1 の増幅(コピー)")
57
3-SAT問題

58
SuyamaのDNAコンピュータ “counting” (Ogihara and Ray)
O(20.4n) molecules for n-variable 3-SAT “dynamic programming” (Suyama) 生成と選択の繰り返し 解の候補の部分的な生成 解の候補の選択 指数オーダーには変わりはないが、 O(20.4n) は O(2n) よりずっと少ない。 固相法 磁気ビーズによるアフィニティ・セパレーション 自動化に適している ⇒ Robot!
 molecules for n-variable 3-SAT. dynamic programming (Suyama) 生成と選択の繰り返し. 解の候補の部分的な生成. 解の候補の選択. 指数オーダーには変わりはないが、 O(20.4n) は O(2n) よりずっと少ない。 固相法. 磁気ビーズによるアフィニティ・セパレーション. 自動化に適している ⇒ Robot!")
59
DNA Computer Robot based on MAGTRATIONTM (Prototype No.1)
")
60
10-variable and 43-clause instance of 3SAT

61
Akira Suyama 東大教授(生物物理)
")
62
ヘアピン・エンジン(SATエンジン) 特平11-165114 Sakamoto et al., Science, May 19, 2000.
DNAのヘアピン構造を利用した選択 ヘアピンの制限酵素切断 exclusive PCR 3-SAT 各節から選んだリテラルから成る一本鎖 DNA 相補的なリテラル = 相補的配列 矛盾したリテラルの選択 ⇒ ヘアピン 6-variable 10-clause 3-SAT Problem SAT計算の本質的部分 = ヘアピン形成 節や変数の数によらないステップ数 Autonomous molecular computation

63
(a∨b∨c)∧(¬d∨e∨¬f)∧ … ∧(¬c∨¬b∨a)∧ ...
ヘアピンの制限酵素切断 Exclusive PCR b ¬b

64
Kensaku Sakamoto 東大&理研(生物化学)
")
65
Adleman-Liptonパラダイムに関する 現在のコンセンサス
電子コンピュータを凌駕するには程遠い。 スケールアップ問題 「分子が計算する」ことの proof of conceptとしては重要。 少なくとも、バイオテクノロジーの ベンチマークとして使うことができる。 さらに、遺伝子計測への応用(Suyama)。
。")
66
Seeman-Winfreeの DNAの自己組織化による計算
3分岐 ... 線形 .... ヘアピン .... DX(ダブルクロスオーバー) 様々なDNA構造分子
 様々なDNA構造分子.")
67
Winfreeの結果 (線形)構造分子によって生成される言語族 =正則言語族 (線形+ヘアピン+3分岐)構造分子によって 生成される言語族
=文脈自由言語族 (線形+DX)構造分子によって =帰納的可算言語族 =チューリング計算可能
構造分子によって. =帰納的可算言語族. =チューリング計算可能.")
68
Winfreeのモデルによる計算過程の例
c = f(a,b) d = g(a,b) c d a b a b Initial Configuration 1次元セルオートマトンの模倣
 d = g(a,b) c. d. a. b. a. b. Initial Configuration. 1次元セルオートマトンの模倣.")
69
Erik Winfree Caltech, Assistant Professor (CS) MacArthur Fellow
Presidential Early Career Award

70
LaBeanたちによるtriple crossover分子

71
分子プログラミング 分子システム設計の計算論的側面 分子の設計 分子反応の設計 分子マシン DNAの場合 = 配列設計
構造 ⇒ 配列(inverse folding) 自己組織化パターンの設計・分子マシンの設計 分子反応の設計 反応条件や操作順序の設計 シミュレーション・ツール 分子マシン 分子プログラミングの現在の目標の一つ
 自己組織化パターンの設計・分子マシンの設計. 分子反応の設計. 反応条件や操作順序の設計. シミュレーション・ツール. 分子マシン. 分子プログラミングの現在の目標の一つ.")
72
配列設計 配列セットの評価 巨大な配列セットの探索 逆問題 ミス・ハイブリダイゼーションの回避 一様な Tm (融解温度)
ハミング距離 エネルギー計算 ⇒ mfold (Zuker), Vienna 一様な Tm (融解温度) 巨大な配列セットの探索 遺伝的アルゴリズム 符号理論 --- Arita’s template method 逆問題 構造 ⇒ 配列(inverse folding) Vienna Group
, Vienna. 一様な Tm (融解温度) 巨大な配列セットの探索. 遺伝的アルゴリズム. 符号理論 --- Arita’s template method. 逆問題. 構造 ⇒ 配列(inverse folding) Vienna Group.")
73
[AT]か [GC] の位置を全配列共通にする
テンプレート法 Arita and Kobayashi, 2002 [AT]か [GC] の位置を全配列共通にする (これをテンプレートと呼ぶ) 例. 011010 より ACCTGA, TGCTCA, TCGACA, etc. → こうすると、全配列の融解温度はほぼ揃う。
![[AT]か [GC] の位置を全配列共通にする](http://slidesplayer.net/slide/14252410/89/images/73/%5BAT%5D%E3%81%8B+%5BGC%5D+%E3%81%AE%E4%BD%8D%E7%BD%AE%E3%82%92%E5%85%A8%E9%85%8D%E5%88%97%E5%85%B1%E9%80%9A%E3%81%AB%E3%81%99%E3%82%8B.jpg "テンプレート法. Arita and Kobayashi, [AT]か [GC] の位置を全配列共通にする. (これをテンプレートと呼ぶ) 例 より. ACCTGA, TGCTCA, TCGACA, etc. → こうすると、全配列の融解温度はほぼ揃う。")
74
どうずらしても、連結した部分を考えても、
ミスマッチを含むテンプレート 上手にテンプレートを選べば、 シフト、リバースの際でも必ずミスマッチをもつ。 例: 110100のとき 110100 110100 110100 110100 110100 110100 110100 どうずらしても、連結した部分を考えても、 ミスマッチは最低2個存在

75
テンプレートの選び方 テンプレート T を、以下のパターンと最低 d 個のミスマッチを持つように選ぶ。 TR TTR 、 TRT
TT、 TRTR 注: TRはTの逆配列。 T=110100なら、001011

76
テンプレートの例 長さ 6 (ミスマッチ 2) 110100 (26個中) 長さ 11 (ミスマッチ 4)
110100 (26個中) 長さ 11 (ミスマッチ 4) , , (211個中) 長さ 23 (ミスマッチ 9) , , (223個中)
 長さ 11 (ミスマッチ 4) , , (211個中) 長さ 23 (ミスマッチ 9) , , (223個中)")
77
DNA配列の設計法 “テンプレート + 誤り訂正符号” 1 1 0 1 0 0 (テンプレート)
誤り訂正符号は何でも利用可。 BCH 符号 Golay 符号 Hamming 符号 など。 A T C A G G (DNA配列) (テンプレート) (任意の符号語)
 (テンプレート) (任意の符号語)")
78
Masanori Arita 東大助教授(バイオインフォマティクス)
")
79
Inverse Folding Vienaグループ McCaskillのアルゴリズムの利用 コスト関数の最小化による配列の探索
Ω : 目的の構造 x: 配列 E(x,Ω): x におけるΩ の自由エネルギー G(x): 配列 x の集団自由エネルギー(McCaskill) p: x におけるΩ の確率
: x におけるΩ の自由エネルギー. G(x): 配列 x の集団自由エネルギー(McCaskill) p: x におけるΩ の確率.")
80
様々な分子マシン

81
SeemanのDNAモーター 2状態のDX分子

82
1) 2) Komiya et al. Whiplash PCR (WPCR) B B A C B A B A C
: stopper sequence B 1) B A C B A B A C 2) This figure shows the scheme of successive transitions. The head A anneals onto the transition table, and then the new state B is polymerized. After the polymerization, the formed hairpin structure is denatured, and the head B is ready for the next transition. Komiya et al.
 B. A. C. B. A. B. A. C. 2) This figure shows the scheme of successive transitions. The head A anneals onto the transition table, and then the new state B is polymerized. After the polymerization, the formed hairpin structure is denatured, and the head B is ready for the next transition. Komiya et al.")
83
3) 4) Whiplash PCR (WPCR) B A B A C B B A C C
The head B, now the current state of the machine, searches and anneals onto the next position in the transition table. The annealing of “B” allows the next state C to be newly polymerized. By iterating this “Annealing, Polymerization, and Denaturation” cycle according to the transition table, DNA molecules perform successive state transitions. B A C 4) C
 C.")
84
Polymerization Stop I explain the basic biochemical reaction composing Whiplash PCR. Each state is encoded by a 15-nucleotide sequence. A sequence at the 3’-end of a DNA molecule works as the head of a state machine, simultaneously representing a current state. A concatenation of two states, termed “state pair”, defines a transition rule. A transition table is represented by a sequence of transition rules. The head anneals onto its complementary sequence in the transition table by forming a hairpin structure intramolecularly, and then a DNA polymerase catalyzes the polymerization of the new state. If the reaction mixture doesn’t contain dTTP, polymerization will stop exactly before the repeated “A”, the stopper sequence. We call this method “Polymerization Stop”.

85
Back-hybridization B A B A C B B A C B A C
Note here that, the new state B can anneal not only onto the next position, but also onto the previous position where B was just polymerized. We call this kind of hairpin formation “back-hybridization”. Back-hybridization does not trigger any polymerization, but may cause a reduction in the efficiency of Whiplash PCR. Distribution of competing alternative hairpin forms and probability of success in transitions should be estimated. Competing Alternative Hairpin Forms

86
Temperature optimization for WPCR
・8 M urea 8% PAGE Komiya, et al. not incubated (℃) This figure shows temperature optimization for Whiplash PCR of 2 rounds on liquid phase with PLATINUM Pfx DNA polymerase. After denaturation at high temperature, transitions were performed under the isothermal condition. Bands that appeared at the highest position are the DNA molecules before transitions. Those at the lowest position are the molecules that finished the 1st round of transitions. And those at the middle position are the molecules that accomplished the 2nd round. The middle band with the highest intensity appeared in the lane of 78℃. Note that, under the thermal cycle schedule like normal PCR, the efficiency decreases. This result is supported by the statistical thermodynamic analysis recently reported in Physical Review E by John A.Rose. Thermal schedule 94℃ for 1 min. ↓ x ℃ for 5 min. x =59.8 ~ 92.2 in 1X Pfx buffer (the composition unknown) 1 mM MgSO4 0.2 mM dATP, dCTP, dGTP 1.5 units Platinum Pfx DNA polymerase
 This figure shows temperature optimization for Whiplash PCR. of 2 rounds on liquid phase with PLATINUM Pfx DNA polymerase. After denaturation at high temperature, transitions were performed under the isothermal condition. Bands that appeared at the highest position are the DNA molecules. before transitions. Those at the lowest position are the molecules that finished the 1st round of transitions. And those at the middle position are the molecules that accomplished the 2nd round. The middle band with the highest intensity appeared in the lane of 78℃. Note that, under the thermal cycle schedule like normal PCR, the efficiency decreases. This result is supported by the statistical thermodynamic analysis recently reported in Physical Review E by John A.Rose. Thermal schedule. 94℃ for 1 min. ↓ x ℃ for 5 min. x =59.8 ~ in 1X Pfx buffer. (the composition unknown) 1 mM MgSO mM dATP, dCTP, dGTP. 1.5 units Platinum Pfx DNA polymerase.")
87
・12% PAGE Successful implementation of transitions Komiya, et al.
( bp ) 155 140 125 110 This picture shows the successful implementation of 8 successive transitions. PCR products are analyzed by 12% polyacrylamide gel electrophoresis. In all lanes from 1 to 8, bands appeared at the positions of the expected length, respectively. The band in the lane i indicates there exist DNA molecules that finished the transition of the ith round and up. We excised the band in the lane 8, and confirmed the implementation of 8 successive transitions by sequencing analysis. 95 80 65 50
 This picture shows the successful implementation of 8 successive transitions. PCR products are analyzed by 12% polyacrylamide gel electrophoresis. In all lanes from 1 to 8, bands appeared at the positions of the expected length, respectively. The band in the lane i indicates there exist DNA molecules that finished the transition of the ith round and up. We excised the band in the lane 8, and confirmed the implementation of 8 successive transitions by sequencing analysis")
88
Ken Komiya 東大(生物化学)
")
89
ShapiroのDNAオートマトン IIS型制限酵素 認識部位 スペーサ a’ 入力の残り <S,a> <S,a>
遷移分子 a’ 入力の残り 入力の残り <S’,a’> 入力文字a’の配列は、各S’に対して<S’,a’>を含んでいる。 遷移分子は、スペーサを調整して、適切な個所で切断する。

90
ShapiroのDNAオートマトン Nature 2001 2入力文字、2状態 FokI a=CTGGCT b=CGCAGC
5’-p…22…GGATGTAC 3’-GGT…22…CCTACATGCCGAp S0,a→S0 5’-p…22…GGATGACGAC 3’-GGT…22…CCTACTGCTGCCGAp S0,a→S1

91
ギネスブック認定 (世界最小のコンピュータ)
")
92
YurkeのDNAピンセット

94
三状態機械

95
Seeman: DNAによるDXの制御

96
System to Test the PX-JX2 Device

97
AFM Evidence for Operation
of the PX-JX2 Device Yan, H., Zhang, X., Shen, Z. & Seeman, N.C. (2002), Nature 415,
, Nature 415,")
98
特定領域「分子プログラミング」計画研究 横森 山下 抽象分子計算系 (解析→設計) 実証分子計算系 (解析→設計→実現) 大内 萩谷 山村
(解析→設計) 自律 近似 分散 実証分子計算系 (解析→設計→実現) 大内 萩谷 山村 高信頼性 自己組織化 進化 関連応用研究 (設計→実現) JST CREST バイオ (遺伝子計測) 情報 ナノテク バイオ (分子進化工学)
 自律. 近似. 分散. 実証分子計算系. (解析→設計→実現) 大内. 萩谷. 山村. 高信頼性. 自己組織化. 進化. 関連応用研究. (設計→実現) JST CREST. バイオ. (遺伝子計測) 情報. ナノテク. バイオ. (分子進化工学)")
99
設計論のための3つの切り口 進化 高信頼性 (設計方法論) (計算過程) 自己組織化 (計算原理) タンパク工学 ドラッグデザイン
バイオテクノロジー 医療応用 ナノテクノロジー 分子エレクトロニクス タンパク工学 ドラッグデザイン 自己組織化 (計算原理) 要するに、分子コンピューティングの応用。
 要するに、分子コンピューティングの応用。")
100
設計論のための3つの切り口 横森 山下 大内 萩谷 山村 進化 高信頼性 (設計方法論) (計算過程) 自己組織化 (計算原理)
バイオテクノロジー 医療応用 ナノテクノロジー 分子エレクトロニクス タンパク工学 ドラッグデザイン 自己組織化 (計算原理) JST CREST 大内 萩谷 山村
 JST CREST. 大内. 萩谷. 山村.")
101
真の分子メモリと分子アドレシング (その心は?)
JST CREST:多相的分子インタラクションに 基づく大容量メモリの構築 多相的分子インタラクション ⇒分子コンピューティングの新たな展開 「真の分子メモリ」の構築 =分子アドレシング技術の確立 光分子コンピューティング(余り話せませんが…) 将来的に自己組織化への応用 自己組織化の光制御
 将来的に自己組織化への応用. 自己組織化の光制御.")
102
真の分子メモリ 分子メモリ 真のメモリ分子 真の分子メモリ 分子や原子の状態によって情報を記憶 分子メモリの本質は、アドレシングにあり。
基本的に可能 いかにして個々の分子に別々の情報を記憶するか。 分子メモリの本質は、アドレシングにあり。 真のメモリ分子 個々の分子が独自のアドレスを持つ。 真の分子メモリ アドレスの異なるメモリ分子の集合体

103
研究の意義 実際的な応用として、 遺伝子情報解析への応用を開発しつつ、 分子アドレシング技術を通して、 分子メモリ構築の基本原理を
確立するとともに、 DNAナノテクノロジーの基本技術として、 将来の情報処理装置の開発に資する。 実際に構築するが、大容量分子メモリそのものではなく、 そのためのアドレシング技術を最終目標。

104
分子アドレシング アドレスを持つ分子の複合体(メモリ)の中から、 指定したアドレスの分子を参照する方法 アドレシングの場
アドレスの数(分子の種類)を多くする --- 大容量 実験操作の手間を減らす --- 高速 アドレシングの場 液相 --- (より大容量・実験操作が簡潔) 固相 --- メモリ分子が固相に固定されている。 (位置情報がアドレスの一部)
を多くする --- 大容量. 実験操作の手間を減らす --- 高速. アドレシングの場. 液相 --- (より大容量・実験操作が簡潔) 固相 --- メモリ分子が固相に固定されている。 (位置情報がアドレスの一部)")
105
液相における分子アドレシング 約600本の正規直交配列セット(現在は約300) 階層メモリ 階層メモリのアドレシング 世界最大のDNAメモリ
100×100×100×100×100×100 = 1006 1060 階層メモリのアドレシング nested PCR WPCR 逐次的ヘアピン開裂 光によるアドレシング アドレシングの高速化

106
Multi-state Molecular Machine
input1 2 1 …… 3 3 input2 1 2 input3 …… 3 3 1 Our goals: Successive state change Input order sensitive …… 2

107
Hairpin-based Machine
階層的アドレシングへの応用

108
Hairpin構造を用いたConformational Addressing (1)
一番端のものにしかアクセスできない。 (赤い数字の部分) あるInput oligomerによってHairpin構造が開かれる と、残されたStem部分は次のヘアピン構造を開く ためにアクセス可能な部分となる。 正しい順番でInput oligomerを加えた場合に のみ、全てのHairpin構造が開かれる。
 あるInput oligomerによってHairpin構造が開かれる. と、残されたStem部分は次のヘアピン構造を開く. ためにアクセス可能な部分となる。 正しい順番でInput oligomerを加えた場合に. のみ、全てのHairpin構造が開かれる。")
109
化学実験による検証 (Single hairpin)
Kameda et al. 電気泳動によってHairpin構造の状態変化を観察した。 (F) (D) (E) (A) Lane data: 1: Hairpin template 2: Input oligomer 3: Probe 4: Hairpin template + Input oligomer 5: Hairpin template + Probe 6: Input oligomer + Probe 7: (Hairpin template + Input oligomer) + Probe 8: (Input oligomer + Probe) + Hairpin template 10% PAGE (non-denaturing gel) 200V const, 35min, SYBR Gold染色
 (D) (E) (A) Lane data: 1: Hairpin template. 2: Input oligomer. 3: Probe. 4: Hairpin template + Input oligomer. 5: Hairpin template + Probe. 6: Input oligomer + Probe. 7: (Hairpin template + Input oligomer) + Probe. 8: (Input oligomer + Probe) + Hairpin template. 10% PAGE (non-denaturing gel) 200V const, 35min, SYBR Gold染色.")
110
化学実験による検証 (2 Hairpin) 実験条件
Kameda et al. Lane 1: 2-Hairpin(0.2 pmol) Lane 2: 2-Hairpin(0.2 pmol) + Input oligomer A (0.5 pmol) Lane 3: 2-Hairpin (0.2 pmol) + Input oligomer B (0.5 pmol) Lane 4: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) Lane 5: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (2.5 pmol) Lane 6: 2-Hairpin (0.2 pmol) + Probe B (0.5 pmol) Lane 7: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Probe B (0.5 pmol) Lane 8: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) Lane 9: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (1.5 pmol) + Probe B (0.5 pmol) Lane 10: Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) Lane 11: Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) 10 % PAGE, 200 V const, 40 min Visualized by FITC or SYBR Gold staining. -反応は全て室温(25℃)で行った。 -それぞれの合成オリゴを10 l (10mM TrisHCl (pH 8.0), 2.5 mM Mg2+)の反応系で混合した。 -各合成オリゴは全て一度に混ぜるのではなく、混合後10分間待ってから次の合成オリゴを添加した。(3種類以上あるLane 4, 5, 7, 8, 9, 11の場合) -Lane ではFITCでラベルしたProbe Bが入っており、FITCを検出することでProbe Bを含む構造のバンドを確認した。
 Lane 2: 2-Hairpin(0.2 pmol) + Input oligomer A (0.5 pmol) Lane 3: 2-Hairpin (0.2 pmol) + Input oligomer B (0.5 pmol) Lane 4: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) Lane 5: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (2.5 pmol) Lane 6: 2-Hairpin (0.2 pmol) + Probe B (0.5 pmol) Lane 7: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Probe B (0.5 pmol) Lane 8: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) Lane 9: 2-Hairpin (0.2 pmol) + Input oligomer A (0.5 pmol) + Input oligomer B (1.5 pmol) + Probe B (0.5 pmol) Lane 10: Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) Lane 11: Input oligomer A (0.5 pmol) + Input oligomer B (0.5 pmol) + Probe B (0.5 pmol) 10 % PAGE, 200 V const, 40 min. Visualized by FITC or SYBR Gold staining. -反応は全て室温(25℃)で行った。 -それぞれの合成オリゴを10 l (10mM TrisHCl (pH 8.0), 2.5 mM Mg2+)の反応系で混合した。 -各合成オリゴは全て一度に混ぜるのではなく、混合後10分間待ってから次の合成オリゴを添加した。(3種類以上あるLane 4, 5, 7, 8, 9, 11の場合) -Lane ではFITCでラベルしたProbe Bが入っており、FITCを検出することでProbe Bを含む構造のバンドを確認した。")
111
化学実験による検証 (2 Hairpin) 実験結果
Kameda et al.

112
固相上の分子アドレシング 位置とアドレスを組み合わせた高密度メモリ トップダウン技術 ⇒ 位置の指定 ボトムアップ技術 さらに、
mm単位の微小連想メモリ トップダウンとボトムアップのハイブリッド トップダウン技術 ⇒ 位置の指定 レーザー(VCSEL) … これは光 AFM陽極酸化 ボトムアップ技術 微小連想メモリ内の分子アドレシング さらに、 微小連想メモリ(アモルファス・メモリ)から 自己組織化分子パターン(結晶化メモリ)へ
 … これは光. AFM陽極酸化. ボトムアップ技術. 微小連想メモリ内の分子アドレシング. さらに、 微小連想メモリ(アモルファス・メモリ)から. 自己組織化分子パターン(結晶化メモリ)へ.")
113
分子アドレシングと自己組織化制御 鶏と卵の関係 すると、例えば、 分子アドレシングにより、 自己組織化パターン上の分子を指定して、
パターンを成長させる。 成長した部分には新しいアドレスが振られる。 すると、例えば、 光によるアドレシング ⇒ 光による自己組織化の制御 トップダウンもボトムアップも

114
まとめ 分子システムによる計算 分子システムを作るための計算 どちらにも、計算機科学の役割は本質的。 将来的に、量子計算に至らずとも、
分子コンピューティング 分子システムを作るための計算 分子プログラミング どちらにも、計算機科学の役割は本質的。 将来的に、量子計算に至らずとも、 分子と量子は切っても切れないだろう。 今後は、化学(特に超分子化学)との 連携が重要。 もの作りのセンス(どちらも役に立たないものを作っている?)
との. 連携が重要。 もの作りのセンス(どちらも役に立たないものを作っている?)")
115
補足 分子コンピューティングの生物への展開は ほとんど述べていない。 遺伝子計測 人工分子進化
インテリジェントDNAチップ 人工分子進化 インテリジェントな(?)タンパク工学 さらに、in-vivo molecular computing インテリジェント・ドラッグ・デリバリ(?)
タンパク工学. さらに、in-vivo molecular computing. インテリジェント・ドラッグ・デリバリ(?)")
Similar presentations


 情報工学科 木村昌臣 篠埜 功.>")
MEMS/NEMS応用展開ビジョンマップ>")


 村田雅之.>")








 15:00ー 場所:葛飾キャンパス研究棟8F第2セミナー室 Speaker:鹿野豊氏>")




 生体高分子解析学講座(第3) スタッフ 教授 新田勝利 助教授 出村誠 助手 相沢智康>")