Download presentation
1
質問・解答例

2
1)CD4陽性T細胞は4種類のエフェクター細胞へ分化するが、それは何が決めているのか?
講義1,2 1)CD4陽性T細胞は4種類のエフェクター細胞へ分化するが、それは何が決めているのか? 2)また、 4種類のエフェクター細胞の機能は何でしょうか? 3)Th1或いはTh2細胞を、どのような細胞を認識し、活性化するのか? 4)Th1或いはTh2細胞は、マクロファージ或いはB細胞をどのように活性化するでしょか? 5) CD8陽性キラー細胞はどのような機構で細胞を殺傷しているのでしょうか?
CD4陽性T細胞は4種類のエフェクター細胞へ分化するが、それは何が決めているのか? 2)また、 4種類のエフェクター細胞の機能は何でしょうか? 3)Th1或いはTh2細胞を、どのような細胞を認識し、活性化するのか? 4)Th1或いはTh2細胞は、マクロファージ或いはB細胞をどのように活性化するでしょか? 5) CD8陽性キラー細胞はどのような機構で細胞を殺傷しているのでしょうか?")
3
・CD4陽性T細胞が、樹状細胞によって抗原提示をされ活性化される際に、近傍に存在するサイトカインの種類によって、決まる。
・IL-12/IFNγが存在すると、Th1細胞へ、IL-4が存在すると、Th2細胞へ、TGFβ+IL-6が存在すると、Th17細胞へ、TGFβのみが存在すると、Tregへ分化する。 2)また、 4種類のエフェクター細胞の機能は何でしょうか? ・Th1細胞は、IFNγを産生し、マクロファージを活性化する。 ・Th2細胞は、IL-4/IL-5を産生し、B細胞を抗体産生細胞へ分化させる。 ・Th17細胞は、炎症性サイトカインIL-17を産生し、上皮細胞・間質細胞に作用し、炎症応答を引き起こす。 ・Treg細胞は、T細胞の活性化を抑制する。
また、 4種類のエフェクター細胞の機能は何でしょうか? ・Th1細胞は、IFNγを産生し、マクロファージを活性化する。 ・Th2細胞は、IL-4/IL-5を産生し、B細胞を抗体産生細胞へ分化させる。 ・Th17細胞は、炎症性サイトカインIL-17を産生し、上皮細胞・間質細胞に作用し、炎症応答を引き起こす。 ・Treg細胞は、T細胞の活性化を抑制する。")
4
活性化された抗原と同じ抗原由来のペプチドをMHCとともに提示するマクロファージ或いはB細胞のみを特異的に認識し、活性化する。
3)Th1或いはTh2細胞を、どのような細胞を認識し、活性化するのか? 活性化された抗原と同じ抗原由来のペプチドをMHCとともに提示するマクロファージ或いはB細胞のみを特異的に認識し、活性化する。 4)Th1或いはTh2細胞は、マクロファージ或いはB細胞をどのように活性化するでしょか? ・Th1細胞はIFN-γを産生するとともに、CD40リガンドを発現して、マクロファージ上のCD40に、結合することによって、殺菌活性を上昇させる。 ・Th2細胞は、IL-4, IL-5を産生するとともに、CD40リガンドを発現して、B細胞上のCD40Lに、結合することによって、抗体産生細胞へ分化誘導する。
Th1或いはTh2細胞を、どのような細胞を認識し、活性化するのか? 活性化された抗原と同じ抗原由来のペプチドをMHCとともに提示するマクロファージ或いはB細胞のみを特異的に認識し、活性化する。 4)Th1或いはTh2細胞は、マクロファージ或いはB細胞をどのように活性化するでしょか? ・Th1細胞はIFN-γを産生するとともに、CD40リガンドを発現して、マクロファージ上のCD40に、結合することによって、殺菌活性を上昇させる。 ・Th2細胞は、IL-4, IL-5を産生するとともに、CD40リガンドを発現して、B細胞上のCD40Lに、結合することによって、抗体産生細胞へ分化誘導する。")
5
5) CD8陽性キラー細胞はどのような機構で細胞を殺傷しているのでしょうか?
・細胞傷害性T細胞は、感染細胞が提示する特異的ペプチドーMHCクラスI複合体を認識すると、傷害顆粒が標的細胞との接着部位に集積し、標的細胞と接触した限局した領域に顆粒の分泌を集中させる。 ・傷害顆粒中のタンパク質分解酵素であるグランザイムが細胞内に入り、標的細胞の細胞質に存在するカスパーゼ3を分解して活性化するとともに、BIDを分解し、ミトコンドリア膜の透過性を上昇させ、チトクロームcを放出されることによってアポトーシスを誘導する。

6
講義3 IL-1~7,10,12、 IFNα、β、γ、TNFα、β、LTs、TGFβ、GM- C- M-CSFの産生細胞と作用について述べなさい。

7
IL-12の作用

8
IL-10 ・自然免疫系の過剰な活性化を抑える ・マクロファージからのIL-1、IL-6、Il-12、TNF-αの産生を抑制する。 ・マクロファージのCD80・CD86の発現を、抑制する。 ・T細胞のMHCクラスII分子や補助受容体の発現を抑制する。 ・Th1細胞からのINF-γ産生を抑制する。

9
講義4 以下の受容体及びそのシグナル伝達について説明しなさい。 IL-3, IL-5, GM-CSF受容体 IL-2, IL-4, IL-7受容体 IFN受容体 TNF受容体 TGF受容体 ケモカイン受容体

10
IL-2, IL-4, IL-7受容体

11
IL-3, IL-5, GM-CSF受容体

12
IFNAR: interferon alpha receptor 1, 2
IFN-α、IFN-β IFNAR: interferon alpha receptor 1, 2 Tyrosine kinase 2

13
IFN-γ IFNGR: interferon gamma receptor 1, 2

14
このJAK-STATシグナル伝達経路の特徴は、核にシグナルが届くまでの最短経路を用いていることである。

15
サイトカイン受容体を介した細胞内シグナルはJAKとSTATによって伝達される
主要サイトカイン受容体においては、その細胞質部分にヤーヌスキナーゼ (Janus kinase; JAK)というチロシンリン酸化酵素が結合していて、受容体が二量体化すると活性化される。 活性化したJAKは受容体のリン酸化チロシン部位に結合した転写因子STAT をリン酸化する。それに伴なって、2分子のSTATが2量体化し、細胞質から核内へ移行してそれぞれのサイトカインに応じた多種の特異的な遺伝子を活性化する。
というチロシンリン酸化酵素が結合していて、受容体が二量体化すると活性化される。 活性化したJAKは受容体のリン酸化チロシン部位に結合した転写因子STAT. をリン酸化する。それに伴なって、2分子のSTATが2量体化し、細胞質から核内へ移行してそれぞれのサイトカインに応じた多種の特異的な遺伝子を活性化する。")
16
TGF (transforming growth factor)受容体
I型受容体とII型受容体 TGFβ I型受容体 II型受容体

17
4量体が形成され、II型受容体はI型受容体をリン酸化する。 I型受容体の酵素活性が上昇し、SMADsをリン酸化
セリン/スレオニンキナーゼ活性 4量体が形成され、II型受容体はI型受容体をリン酸化する。 I型受容体の酵素活性が上昇し、SMADsをリン酸化 タイプII受容体にTGFβが結合する。 増殖を抑制する働きがある。

18
1)B細胞の分化段階とH鎖遺伝子とL鎖遺伝子の再編成、発現についてまとめる。
講義6,7 1)B細胞の分化段階とH鎖遺伝子とL鎖遺伝子の再編成、発現についてまとめる。 2)B細胞の免疫寛容の仕組み 3)抗原と出会うと起こるB細胞抗原受容体の変化について 4)ヘルパーT細胞存在下で起こるクラススイッチの仕組み 5)B細胞の分化に行う生体内移動について
B細胞の分化段階とH鎖遺伝子とL鎖遺伝子の再編成、発現についてまとめる。 2)B細胞の免疫寛容の仕組み. 3)抗原と出会うと起こるB細胞抗原受容体の変化について. 4)ヘルパーT細胞存在下で起こるクラススイッチの仕組み. 5)B細胞の分化に行う生体内移動について.")
19
1) 骨髄内 H鎖遺伝子 L鎖遺伝子 生殖細胞系列 幹細胞 D-J 再編成 V-DJ 再編成 早期プロB細胞 後期プロB細胞
膜型mH鎖の産生 幹細胞 早期プロB細胞 後期プロB細胞 大型プレB細胞 小型プレB細胞 未熟B細胞 V-J 再編成 再編成されたVJ H鎖遺伝子 L鎖遺伝子 骨髄内 再編成されたVDJ 選択的スプライシング 膜型mH, dH鎖の産生 再編成されたVJ 成熟B細胞

20
2) トレランス(自己寛容性)の誘導 自己抗原を認識すると、 ・受容体編集(Receptor editing:レセプターエディティング) ・アポトーシスによるクローン消失 ・アナジー
 トレランス(自己寛容性)の誘導 自己抗原を認識すると、 ・受容体編集(Receptor editing:レセプターエディティング) ・アポトーシスによるクローン消失 ・アナジー")
21
受容体編集(レセプターエディティング) 多価の自己抗原に強く架橋すると細胞表面のIgM発現が低下しRAG遺伝子の発現が保たれ、L鎖遺伝子の再編成が繰り返し行われる。 クローン消失 (clonal deletion) 救済され、成熟B細胞へ分化する。

22
アナジー(不応答性) 自己反応性のアナジー状態のB細胞は、CXCR5の発現が低く、CXCL13(濾胞樹状細胞が出している)に対する反応性が悪いため、たどり着けない。
 自己反応性のアナジー状態のB細胞は、CXCR5の発現が低く、CXCL13(濾胞樹状細胞が出している)に対する反応性が悪いため、たどり着けない。")
23
末梢性自己寛容 特定臓器のみにしか発現していない自己抗原がたくさんある。 末梢で自己抗原に反応するB細胞が活性化しない仕組みが存在する。
外来抗原に応答するB細胞の場合はそれに相当するヘルパーT細胞がT細胞領域に存在するのに対して、自己抗原に応答するB細胞の場合はそのようなヘルパーT細胞は存在しない。 T細胞と相互作用するとCXCR5が発現するので、濾胞に入ることができる。T細胞領域からB細胞濾胞へ移行することができない。B細胞濾胞へ移動できないB細胞は、生存シグナルを受けることができないのですぐにアポトーシスを起こして消失する。

24
3)抗原と出会うと起こるB細胞抗原受容体の変化について
1)膜型から分泌型免疫グロブリン(抗体)への変化 2)抗原特異性は変化しないが、異なったC領域の免疫グロブリンが産生される。クラススイッチ;アイソタイプスイッチ 3)V領域の配列が、体細胞高頻度変異 (somatic hypermutation)によりさらに多様化し、抗原により高い親和性をもった抗体を産生するB細胞が形質細胞へ分化しやすくなる。親和性成熟(affinity maturation) 4)記憶B細胞(メモリB細胞)の生成。
膜型から分泌型免疫グロブリン(抗体)への変化. 2)抗原特異性は変化しないが、異なったC領域の免疫グロブリンが産生される。クラススイッチ;アイソタイプスイッチ. 3)V領域の配列が、体細胞高頻度変異 (somatic hypermutation)によりさらに多様化し、抗原により高い親和性をもった抗体を産生するB細胞が形質細胞へ分化しやすくなる。親和性成熟(affinity maturation) 4)記憶B細胞(メモリB細胞)の生成。")
25
4)ヘルパーT細胞存在下で起こるクラススイッチの仕組み
クラススイッチは「遺伝子組み換え」(DNA再編成)によって起こる。 H鎖の各C遺伝子(Cm, Cd, Cg3, Cg1,Ca1,Cg2, Cg4, Ce, Ca2)の5’側にはDNAの反復配列(S領域)がある。S領域間で組み換えが起こり、その間のDNAは除去され、IgM, IgD以外のアイソタイプが発現する。 まず最初に、mスイッチ領域(Sm)が他のS領域と結合することによって起こり、さらに、他のアイソタイプへスイッチは、このmスイッチが生じたところから起こる。
によって起こる。 H鎖の各C遺伝子(Cm, Cd, Cg3, Cg1,Ca1,Cg2, Cg4, Ce, Ca2)の5’側にはDNAの反復配列(S領域)がある。S領域間で組み換えが起こり、その間のDNAは除去され、IgM, IgD以外のアイソタイプが発現する。 まず最初に、mスイッチ領域(Sm)が他のS領域と結合することによって起こり、さらに、他のアイソタイプへスイッチは、このmスイッチが生じたところから起こる。")
26
AID; activation-induced cytidine deaminase, 活性化誘導シチジンデアミナーゼ
体細胞突然変異を誘導する酵素 AID; activation-induced cytidine deaminase, 活性化誘導シチジンデアミナーゼ DNAやRNAを構成するシトシンをウラシルに変換する酵素

27
5)B細胞の分化に行う生体内移動について 1)B細胞は、骨髄内で幹細胞から膜型免疫グロブリン(B細胞抗原受容体)をもつ成熟B細胞へ分化する。 2)成熟B細胞は、血流を介して2次リンパ組織を再循環しながら全身を移動する。 3)2次リンパ組織(リンパ節、脾臓、パイエル板)で抗原に遭遇すると、活性化され、同じ抗原に特異的なヘルパーT細胞と胚中心を作り、そこで抗体産生細胞(形質細胞)へ分化する。 4)形質細胞は、骨髄へ移動し抗体を産生する。
2次リンパ組織(リンパ節、脾臓、パイエル板)で抗原に遭遇すると、活性化され、同じ抗原に特異的なヘルパーT細胞と胚中心を作り、そこで抗体産生細胞(形質細胞)へ分化する。 4)形質細胞は、骨髄へ移動し抗体を産生する。")
28
1)感染防御において産生される抗体のサブクラスの推移について説明しなさい。 2)TI抗原の化学的性質と抗体応答の特徴について説明しなさい。
講義8 1)感染防御において産生される抗体のサブクラスの推移について説明しなさい。 2)TI抗原の化学的性質と抗体応答の特徴について説明しなさい。 3)B細胞抗原受容体のITAMモチーフについて説明しなさい。 4)B細胞抗原受容体の二つの役割とは? 5)高親和性で、エフェクター機能を有するサブクラスの抗体産生するB細胞はどのような環境でできるのか? 6)高親和性抗体が生成される仕組みを説明しなさい。
感染防御において産生される抗体のサブクラスの推移について説明しなさい。 2)TI抗原の化学的性質と抗体応答の特徴について説明しなさい。 3)B細胞抗原受容体のITAMモチーフについて説明しなさい。 4)B細胞抗原受容体の二つの役割とは? 5)高親和性で、エフェクター機能を有するサブクラスの抗体産生するB細胞はどのような環境でできるのか? 6)高親和性抗体が生成される仕組みを説明しなさい。")
29
1)感染防御において産生される抗体のサブクラスの推移について
ヘルパーT細胞の補助なしで産生される抗体や感染初期に産生される抗体は低親和性のIgMである。(免疫記憶も長期にわたる感染防御もできない。初期の感染を低いレベルに抑える。) 2次リンパ組織の胚中心で、ヘルパーT細胞との相互作用によって、クラススイッチや体細胞高頻度変異、親和性成熟を起こし、高親和性のIgG, IgAを産生する形質細胞が生じる。
 2次リンパ組織の胚中心で、ヘルパーT細胞との相互作用によって、クラススイッチや体細胞高頻度変異、親和性成熟を起こし、高親和性のIgG, IgAを産生する形質細胞が生じる。")
30
2)TI抗原の化学的性質と抗体応答の特徴 TI抗原とは、T細胞の補助なしで抗体産生を誘導する抗原。リポ多糖類、ペプチドグリカンなど反復する糖質エピトープを有する。 産生される抗体は、IgMのみで体細胞高頻度変異も起こらないので、抗体の抗原に対する親和性は亢進しないし、免疫記憶も誘導されない。
TI抗原の化学的性質と抗体応答の特徴 TI抗原とは、T細胞の補助なしで抗体産生を誘導する抗原。リポ多糖類、ペプチドグリカンなど反復する糖質エピトープを有する。 産生される抗体は、IgMのみで体細胞高頻度変異も起こらないので、抗体の抗原に対する親和性は亢進しないし、免疫記憶も誘導されない。")
31
3)B細胞抗原受容体のITAMモチーフについて説明しなさい。
ITAMモチーフ(免疫受容体チロシン活性化モチーフ:D/E)X 7(D/E)X2(Y)X2(L/I)X7(Y)X2(L/I))は、チロシン残基を二つ含みシグナル伝達に関与 するモチーフで、Igα, Igβの細胞質部分に存在する。 細胞表面IgMに、特異抗原結合すると、Src型チロシンキナーゼ(Blk, Fyn, Lyn)が活性化され、IgβのITAMのチロシン残基をリン酸化する。 Sykが、リン酸化されたIgβのITAMに結合する。Syk分子は近接して、相互にリン酸化し合って活性化される。最終的には核まで伝達されて、B細胞活性化に必要な遺伝子発現を誘導する。
X 7(D/E)X2(Y)X2(L/I)X7(Y)X2(L/I))は、チロシン残基を二つ含みシグナル伝達に関与 するモチーフで、Igα, Igβの細胞質部分に存在する。 細胞表面IgMに、特異抗原結合すると、Src型チロシンキナーゼ(Blk, Fyn, Lyn)が活性化され、IgβのITAMのチロシン残基をリン酸化する。 Sykが、リン酸化されたIgβのITAMに結合する。Syk分子は近接して、相互にリン酸化し合って活性化される。最終的には核まで伝達されて、B細胞活性化に必要な遺伝子発現を誘導する。")
32
4)B細胞抗原受容体の二つの役割とは? 1) B細胞抗原受容体は、抗原と結合してシグナルを細胞内へ伝達する。このとき、B細胞補助受容体からのシグナルが必要である。 2)抗原受容体を介して、抗原を取込み、処理しMHCクラスII-ペプチド複合体をヘルパーT細胞へ提示する。B細胞の増殖・分化には、ヘルパーT細胞からの第2シグナル(サイトカイン、CD40L)が必要である。
抗原受容体を介して、抗原を取込み、処理しMHCクラスII-ペプチド複合体をヘルパーT細胞へ提示する。B細胞の増殖・分化には、ヘルパーT細胞からの第2シグナル(サイトカイン、CD40L)が必要である。")
33
5)高親和性で、エフェクター機能を有するサブクラスの抗体産生するB細胞はどのような環境でできるのか?
血中を循環するナイーブB細胞は、高内皮性小静脈(HEV)経由で血中からリンパ節のT細胞領域に入る。同じ抗原に特異的なヘルパーTh2細胞に出会うと、結合し、増殖する。活性化B細胞とTh2細胞とから構成される一次反応巣を形成する。活性化B細胞は、ヘルパーTh2細胞へ密着したまま、一次リンパ瀘胞へ移動し、胚中心(germinal center)を作る。胚中心は、親和性成熟とクラススイッチに適した特殊化した微小環境を構成する。
経由で血中からリンパ節のT細胞領域に入る。同じ抗原に特異的なヘルパーTh2細胞に出会うと、結合し、増殖する。活性化B細胞とTh2細胞とから構成される一次反応巣を形成する。活性化B細胞は、ヘルパーTh2細胞へ密着したまま、一次リンパ瀘胞へ移動し、胚中心(germinal center)を作る。胚中心は、親和性成熟とクラススイッチに適した特殊化した微小環境を構成する。")
34
6)高親和性抗体が生成される仕組みを説明しなさい。
胚中心の暗領域で、T細胞由来のサイトカインによって、分裂・増殖している中心芽細胞で、B細胞抗原受容体の体細胞高頻度変異が起こる。抗原受容体に変異を起こした中心細胞が生じる。中心細胞は、親和性の高いものから低いものまで、幅の広い親和性をもった膜型免疫グロブリンを発現している。 体細胞高頻度変異した中心細胞はお互い競合し、瀘胞樹状細胞上の抗原と抗原特異的ヘルパーT細胞を奪い合う。高親和性抗原受容体をもった中心細胞ほど、濾胞樹状細胞より、抗原を取り込むことが出来るので、ペプチド-MHCクラスII複合体を、ヘルパーT細胞へ提示でき、ヘルパーT細胞からサイトカインやCD40Lの刺激を受け取ることができ、アポトーシスを免れ、抗体産生細胞へ分化する。

35
2) Fc領域の異なる抗体とその受容体の種類と働き
講義9 1) 抗体の二つの働き 2) Fc領域の異なる抗体とその受容体の種類と働き
 抗体の二つの働き. 2) Fc領域の異なる抗体とその受容体の種類と働き.")
36
・抗体の働きは『中和』 『オプソニン化』。
1)抗体の二つの働き ・抗体の働きは『中和』 『オプソニン化』。 ・クラススイッチにより、Fc領域が違う抗体ができる。Fc領域の機能は体の部位に抗体を運搬することと抗体に結合した抗原とそれを破壊する分子や細胞を結び付ける。
抗体の二つの働き. ・抗体の働きは『中和』 『オプソニン化』。 ・クラススイッチにより、Fc領域が違う抗体ができる。Fc領域の機能は体の部位に抗体を運搬することと抗体に結合した抗原とそれを破壊する分子や細胞を結び付ける。")
37
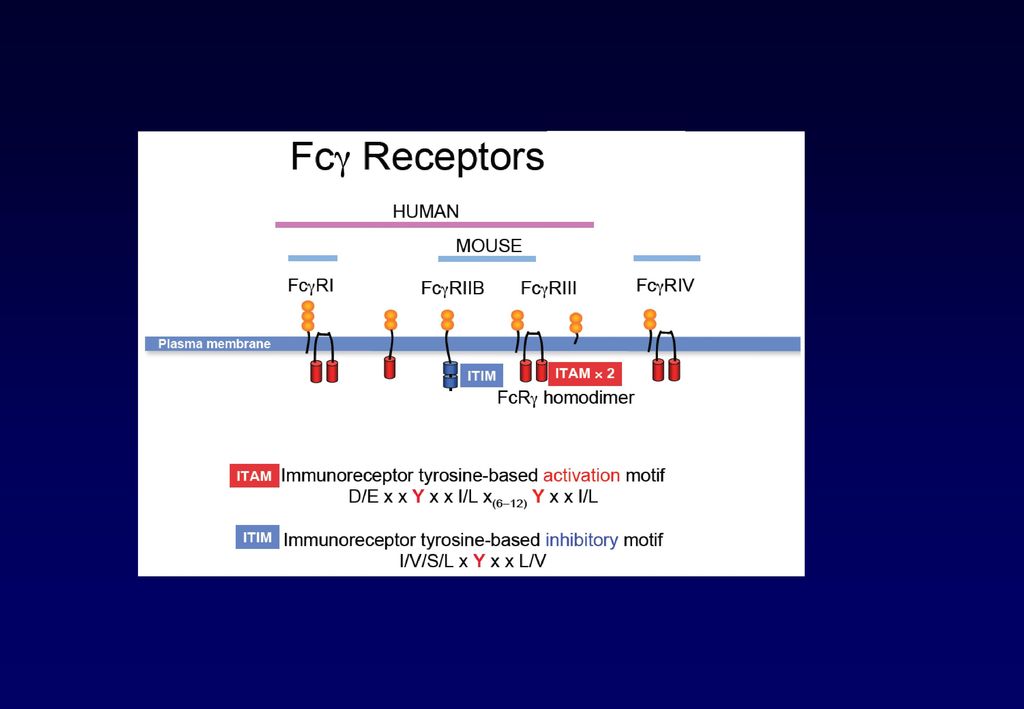
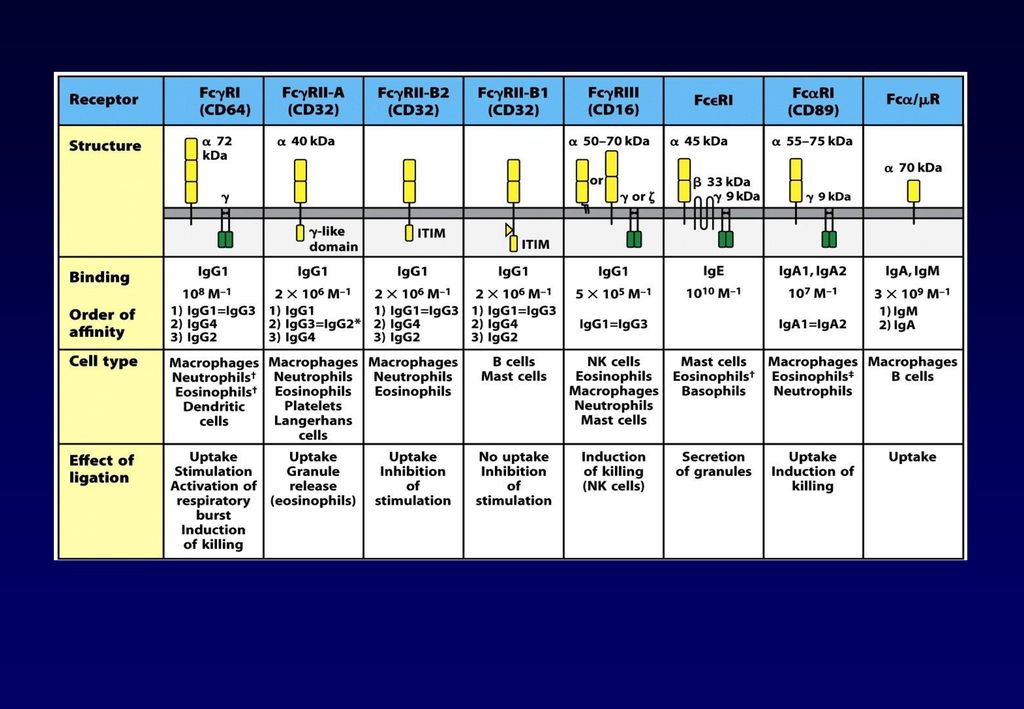
・マクロファージ/好中球などの貪食細胞は、主にIgGのFc領域に結合するFcγ受容体は発現し、IgGでオプソニン化された病原体を破壊する。
・マスト細胞、好塩基球、好酸球上には、IgEが高親和性のFcε受容体にもともと結合しており、抗原に結合すると、脱顆粒し、炎症メディエーターを放出し、寄生虫を物理的に排除する。

40
講義10 1)補体活性化の3つの経路? 2)補体の主な3つの機能? 3)C3の分解と働き?
補体活性化の3つの経路? 2)補体の主な3つの機能? 3)C3の分解と働き?")
41
1)補体系の活性化経路 1)古典経路 IgM或はIgG抗体が微生物表面に結合することによって誘導される。 2)レクチン経路 細菌壁の糖鎖に結合する血漿中のマンノース結合レクチンによって開始される。 3)第二経路 ある種の細菌の表面を構成する物理的な局所環境によって、非特異的にC3が、結合し活性化する。
第二経路. ある種の細菌の表面を構成する物理的な局所環境によって、非特異的にC3が、結合し活性化する。")
42
・C3bが微生物に結合する(オプソニン化)と、補体レセプターを介する食細胞のファゴサイトーシスが促進される。
2)補体の主な3つの機能 ・C3bが微生物に結合する(オプソニン化)と、補体レセプターを介する食細胞のファゴサイトーシスが促進される。 ・C3bが微生物に結合すると、補体の後期構成要素が活性化によって、膜攻撃複合体が形成され、浸透圧溶解を起こす。 ・C5a, C3a, C4aによっては白血球の動員及び活性化が起こり、炎症が誘導され、微生物は破壊される。
補体の主な3つの機能. ・C3bが微生物に結合する(オプソニン化)と、補体レセプターを介する食細胞のファゴサイトーシスが促進される。 ・C3bが微生物に結合すると、補体の後期構成要素が活性化によって、膜攻撃複合体が形成され、浸透圧溶解を起こす。 ・C5a, C3a, C4aによっては白血球の動員及び活性化が起こり、炎症が誘導され、微生物は破壊される。")
43
C3bはさらに蛋白分解酵素により、iC3b, C3dg, C3dに分解される。
C4b2a(b)複合体及びC3bBb複合体は、C3転換酵素で、補体成分C3をC3aとC3bに分解する。 C3bは病原体の表面に結合し異物としてマーキングする。 C3bはさらに蛋白分解酵素により、iC3b, C3dg, C3dに分解される。 C3b → iC3b + C3f → C3dg + C3c → C3d + C3g C3b,iC3b,C3dg,C3dは補体受容体に認識される。 補体受容体 CR1 C3a CR2 CR3 CR4
複合体及びC3bBb複合体は、C3転換酵素で、補体成分C3をC3aとC3bに分解する。 C3bは病原体の表面に結合し異物としてマーキングする。 C3bはさらに蛋白分解酵素により、iC3b, C3dg, C3dに分解される。 C3b → iC3b + C3f → C3dg + C3c → C3d + C3g. C3b,iC3b,C3dg,C3dは補体受容体に認識される。 補体受容体. CR1. C3a. CR2. CR3. CR4.")
44
まとめ ・自然免疫の推移? ・貪食受容体について説明しなさい。 ・TLR受容体の種類とサイトカイン産生するシグナル伝達経路について説明しなさい。 ・自然免疫が、獲得免疫に果たす役割と、獲得免疫へ与える影響? ・NK/NKT細胞の認識機構と機能について説明しなさい。

45
自然免疫の推移? 1)病原体が上皮細胞の障壁をすり抜けて、組織に侵入すると、最初に補体が、第2経路によって活性化される。 2)マクロファージは、病原微生物に共通して存在する特有の分子構造を認識する。これらの受容体はパターン認識受容体(PRR, pattern recognition receptor)と呼ばれ、食作用に関与する受容体(マンノース受容体、スカベンジャー受容体、補体受容体)とシグナル型受容体TLR(Toll-like receptor)などがある。 3)マクロファージは、貪食受容体を介して病原体を取り込む。また、Toll様受容体(TLR)を介して、種々の炎症性サイトカインを産生し、好中球の感染組織への浸潤や、発熱などの全身性の反応を引き起こす。
マクロファージは、病原微生物に共通して存在する特有の分子構造を認識する。これらの受容体はパターン認識受容体(PRR, pattern recognition receptor)と呼ばれ、食作用に関与する受容体(マンノース受容体、スカベンジャー受容体、補体受容体)とシグナル型受容体TLR(Toll-like receptor)などがある。 3)マクロファージは、貪食受容体を介して病原体を取り込む。また、Toll様受容体(TLR)を介して、種々の炎症性サイトカインを産生し、好中球の感染組織への浸潤や、発熱などの全身性の反応を引き起こす。")
46
貪食受容体について説明しなさい。 CR1 CR3 CR4 マクロファージは、第2経路による補体活性化を通して、病原体表面にC3b断片が結合すると、補体受容体CR1を介して、より効率的に、その病原体を貪食する。 C3bがI因子で分解され、iC3bへと分解されると、補体受容体CR3やCR4によって認識される。 マンノース受容体 レクチンは、ヒトの細胞にはない病原体特有の糖鎖構造、立体構造に結合する。HIVなどの特定のウイルスや細菌の高マンノース糖鎖に結合する。 スカベンジャー受容体 変性LDL(酸化LDLなど)を認識して取り込む。
を認識して取り込む。")
47
TLR受容体の種類とサイトカイン産生するシグナル伝達経路について説明しなさい。

48
Nature Immunology, 2011 TLR4 signaling activates the canonical IKK complex via MyD88-dependent and TRIF-dependent mechanisms. MyD88 assembles complexes that contain IRAK kinases together with TRAF6, TAB2, TAB3 and TAK1. TRIF can directly recruit TRAF6 and recruit TAB2, TAB3 and TAK1. The active NF-κB pathway subsequently induces expression of the alternative MyD88 splice product MyD88s, the kinase-inactive IRAK family member IRAK-M and the negative regulatory adaptor molecule SARM. These dominant-negative factors presumably affect the stability of the IKK-activating complexes.

49
自然免疫が、獲得免疫に果たす役割と、獲得免疫へ与える影響?
・感染組織から近傍のリンパ組織へ、コスティムレーター(B7)を発現した樹状細胞によって抗原情報が伝達され、ナイーブT細胞が活性化され獲得免疫が開始する。 ・自然免疫に関与する細胞により産生されるサイトカインは、CD4T細胞が、Th1細胞或はTh2細胞へ分化すのかを決定する。
を発現した樹状細胞によって抗原情報が伝達され、ナイーブT細胞が活性化され獲得免疫が開始する。 ・自然免疫に関与する細胞により産生されるサイトカインは、CD4T細胞が、Th1細胞或はTh2細胞へ分化すのかを決定する。")
50
NK/NKT細胞の認識機構と機能について説明しなさい。
・ウイルス感染細胞・腫瘍細胞にアポトーシスを誘導し、破壊する。 ・FN-γ を産生してマクロファージを活性化する。

51
NK細胞は、自己MHCと決定づけられるものの欠損により、攻撃が引き起こされる。『失われた自己』を認識している。

52
T細胞と同様にTCRを介して抗原を認識しますが、その多様性は乏しく (invariant)、マウスでは、α鎖はVα14、β鎖はVβ8
T細胞と同様にTCRを介して抗原を認識しますが、その多様性は乏しく (invariant)、マウスでは、α鎖はVα14、β鎖はVβ8.2、Vβ7、Vβ2、ヒトではα鎖はVα24、β鎖はVβ11だけを発現している スフィンゴ糖脂質:α-Galactosylceramide (α-GalCer)を認識している。
、マウスでは、α鎖はVα14、β鎖はVβ8.2、Vβ7、Vβ2、ヒトではα鎖はVα24、β鎖はVβ11だけを発現している. スフィンゴ糖脂質:α-Galactosylceramide (α-GalCer)を認識している。")
53
1) I型アレルギー(即時型アレルギー)の原因と推移について
2) 近年、アレルギーが増加した要因?
 近年、アレルギーが増加した要因?")
54
1)I型アレルギー(即時型アレルギー)の原因
アレルゲンとの接触で、IgEを介するマスト細胞,好酸球の脱顆粒が起き、炎症メディエーターやサイトカインが放出され、アレルギー症状が起きる。 IgEにアレルゲンが結合すると、FcεRIが架橋され、シグナルが細胞内に伝わり、炎症物質が放出、生成される。 主要なメディエーターは、顆粒から放出される血管作動性アミン(ヒスタミンなど)とプロテアーゼ、アラキドン酸の代謝産物、およびサイトカインである。
とプロテアーゼ、アラキドン酸の代謝産物、およびサイトカインである。")
55
1)I型アレルギー(即時型アレルギー)の推移
I 型アレルギーには、「即時型反応」とその後に続く「遅発型反応」がある。 ・「即時型反応」では、アレルゲンにマスト細胞(肥満細胞)が反応する。マスト細胞から放出されるケミカルメディエーターやサイトカイン、ケモカインの作用によって、循環中のTh2ヘルパーT細胞、好酸球がアレルゲン侵入組織に集まって、さらに強力で有害な反応を起こす。 ・「遅発型反応」の主役は好酸球で、慢性的なアレルギー炎症の原因。 アレルゲンがヘルパーTh2細胞を活性化すると、IL5などのサイトカインが産生され、骨髄での好酸球生成と末梢血中への放流が上昇する。 好酸球は、マスト細胞が産生するケモカイン(Eotaxinなど)やECF-Aの刺激で、組織へ移動し、活性化される。好酸球は強い毒性をもつプロスタグランジン、ロイコトリエン、MBPを産生し寄生虫に対すると同様に宿主を傷害する。
が反応する。マスト細胞から放出されるケミカルメディエーターやサイトカイン、ケモカインの作用によって、循環中のTh2ヘルパーT細胞、好酸球がアレルゲン侵入組織に集まって、さらに強力で有害な反応を起こす。 ・「遅発型反応」の主役は好酸球で、慢性的なアレルギー炎症の原因。 アレルゲンがヘルパーTh2細胞を活性化すると、IL5などのサイトカインが産生され、骨髄での好酸球生成と末梢血中への放流が上昇する。 好酸球は、マスト細胞が産生するケモカイン(Eotaxinなど)やECF-Aの刺激で、組織へ移動し、活性化される。好酸球は強い毒性をもつプロスタグランジン、ロイコトリエン、MBPを産生し寄生虫に対すると同様に宿主を傷害する。")
56
2) 近年、アレルギーが増加した要因? ・アレルゲンはTh2型の応答を起こし、IgE産生を誘導する。ほとんどは空中に浮遊する限られた種類の蛋白質抗原で、微量に侵入するものである。小さな可溶性分子。 ・近年、アレルギーが増加した要因 1)アレルゲンの量が増えた。 2) 幼児期の感染症が激減したため、Th2応答が優性となった。 3)大気汚染物質の増加。
アレルゲンの量が増えた。 2) 幼児期の感染症が激減したため、Th2応答が優性となった。 3)大気汚染物質の増加。")


は、腹足綱後鰓類の無楯類 (Anapsidea, Aplysiomorpha) に属する軟体動物の総称。 より.>")
 英国のジェンナーは、牛飼いの女性から牛 痘に罹ったものが天然痘にならないことを 聞いて、牛痘の接種による天然痘の予防法 を思いついた。牛痘の膿を少年に接種し、 その後真性の凍瘡を接種しても発病を免れ たことから、予防接種の有効性が証明され た。(1796年)>")
 法. 3.ミニブタでの遺伝子改変 (gene targeting) 法. -マウスと異種移植用動物での gene targeting 法の違い- 4.異種移植用遺伝子改変ミニブタ開発における現在の問題点と解決の方.>")





:16-23. Resveratrol inhibition of human keratinocyte proliferation via SIRT1/ARNT/ERK dependent downregulation of AQP3 レスベラトロールによるヒトケラチノサイトの増殖阻害は.>")



.>")





