Download presentation
1
ゲノム・遺伝子・DNA 染色体=タンパク質(ヒストン)+DNA
細胞分裂時の染色体(青)と紡錘糸(緑Microtubule)
と紡錘糸(緑Microtubule)")
7
遺伝子とDNA 遺伝子:細胞から細胞へ、親から子へ受け継がれ、肌の色や性格など特定の性質(分子レベルで見ると主に蛋白質の一次構造=アミノ酸の並び方)を規定するもの(抽象的な概念) DNAはその物質的実体、直線あるいは環状分子で、遺伝子を構成する領域が並んでいる(上のイメージ) 染色体とは、そのDNAを含む目に(顕微鏡で)見える構造
見える構造.")
8
ヒトの23対の染色体

9
X染色体と Y染色体

10
染色体

12
DNAの折りたたみ Nucleosomes-H1 linker histone
Figure Model for the packing of chromatin and the chromosome scaffold in metaphase chromosomes. In interphase chromosomes, long stretches of 30-nm chromatin loop out from extended scaffolds. In metaphase chromosomes, the scaffold is folded into a helix and further packed into a highly compacted structure, whose precise geometry has not been determined. Fig. 2. Models for the 30-nm fiber. The figure shows a schematic representation of two models that have been proposed to explain the 30-nm fiber, as discussed in the text. The electron micrographs of the 11-nm fiber and solenoid were adapted and modified from Molecular Biology of the Cell (B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts, P. Walter, Garland Science Publishing, London, ed. 4, 2002). The zig-zag electron micrograph was adapted and modified from J. Cell Biol. 83, 403 (1979). Two concentrations of TEA-Cl (trieathanolamine-HCl) are shown, 1 and 5 mM, to emphasize the importance of the salt concentration in the compaction of the nucleosomal arrays. Changing faces of chromatin.htm
. The zig-zag electron micrograph was adapted and modified from J. Cell Biol. 83, 403 (1979). Two concentrations of TEA-Cl (trieathanolamine-HCl) are shown, 1 and 5 mM, to emphasize the importance of the salt concentration in the compaction of the nucleosomal arrays. Changing faces of chromatin.htm.")
13
Intron(余計な切り取られるところ)とExon(タンパク質に翻訳されるところ)
ヒトの染色体の構成

15
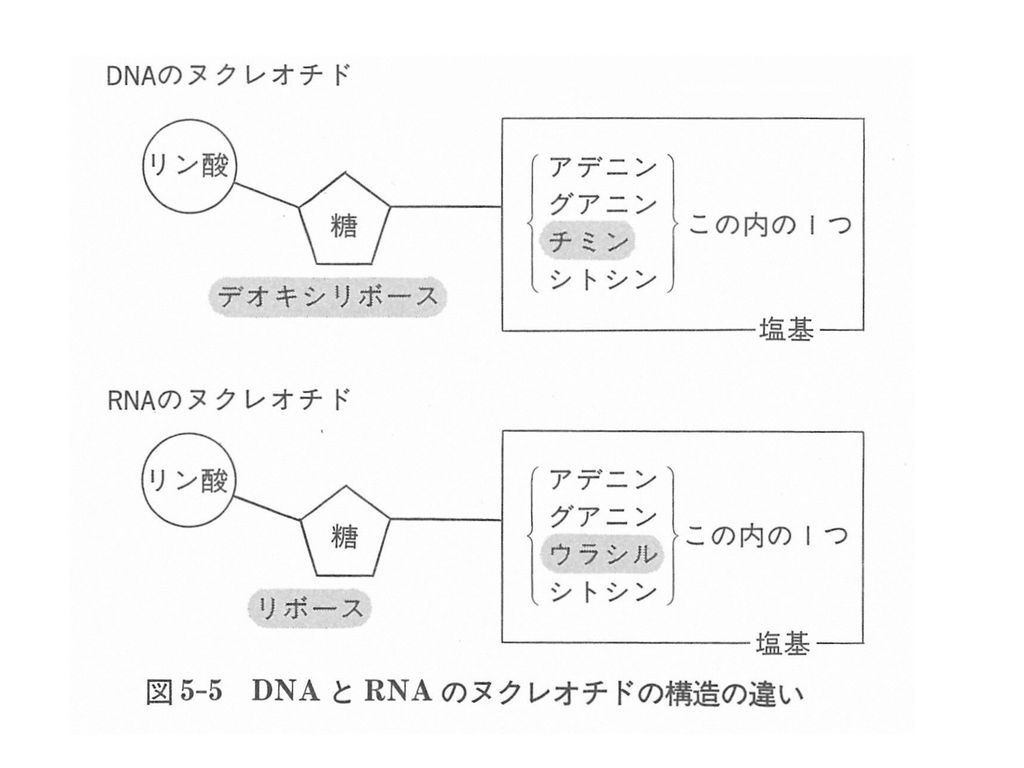
DNAとRNA

16
4(5)つの塩基 RNA
つの塩基 RNA")
17
AとT(U) GとCが水素結合する 塩基対形成Base pairing
 GとCが水素結合する 塩基対形成Base pairing")
19
二重らせん double helix 水素結合の重要性
核酸 染色体のDNAは、2本の高分子が対になっていて、2重らせん構造をしています。対を作っているのは、アデニン(A)、チミン(T)、グアニン(G)、シトシン(C)の核酸塩基とよばれるものです。水の水素結合は結構でたらめですが、核酸塩基の水素結合は決まった組み合わせで作られます。アデニンはチミンと、グアニンはシトシンと図のような水素結合しています。
、チミン(T)、グアニン(G)、シトシン(C)の核酸塩基とよばれるものです。水の水素結合は結構でたらめですが、核酸塩基の水素結合は決まった組み合わせで作られます。アデニンはチミンと、グアニンはシトシンと図のような水素結合しています。")
20
同じく水素結合で保持されているが、DNAは塩基を内側に(塩基間の水素結合)、蛋白質のα-Helix は,主鎖間の水素結合で成り立ち,アミノ酸側鎖を外側に出している
、蛋白質のα-Helix は,主鎖間の水素結合で成り立ち,アミノ酸側鎖を外側に出している")
21
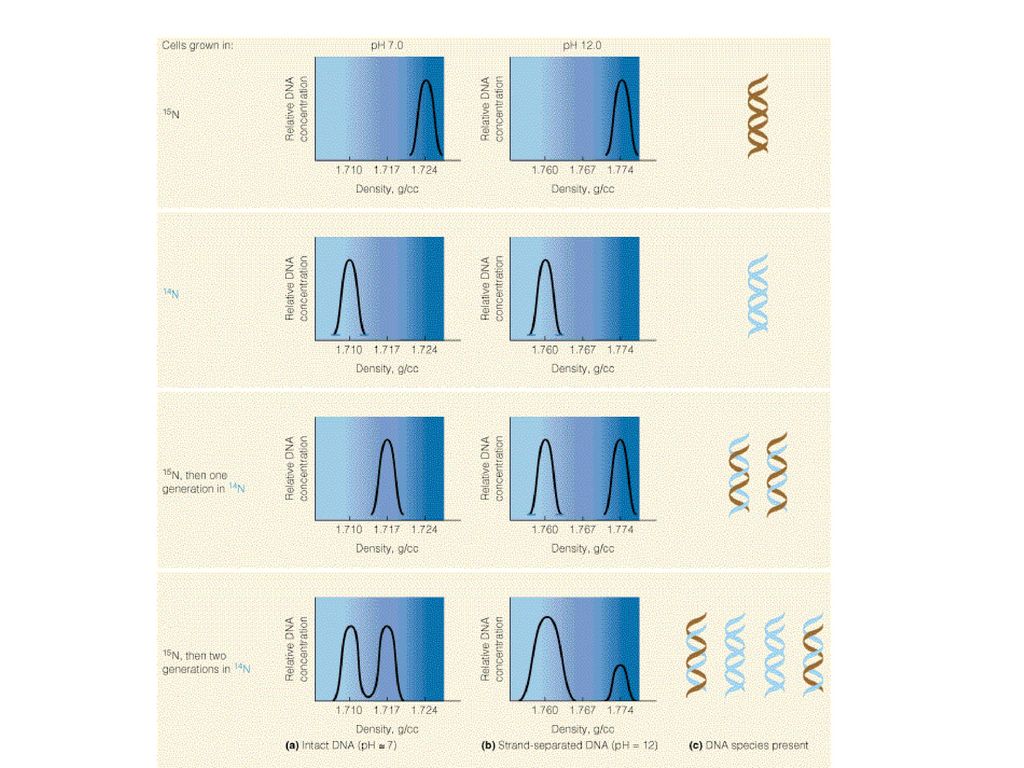
DNAの複製 Replication 半保存的複製

23
DNA複製と細胞分裂 大腸菌のDNAは環状

24
複製開始が頻繁だとこんな感じ 半保存的複製と分かる

25
遺伝情報発現の Central Dogma DNA 複製 Replication 転写 Transcription RNA
翻訳 Translation 蛋白質

26
転写 Transcription =mRNA合成

27
転写

28
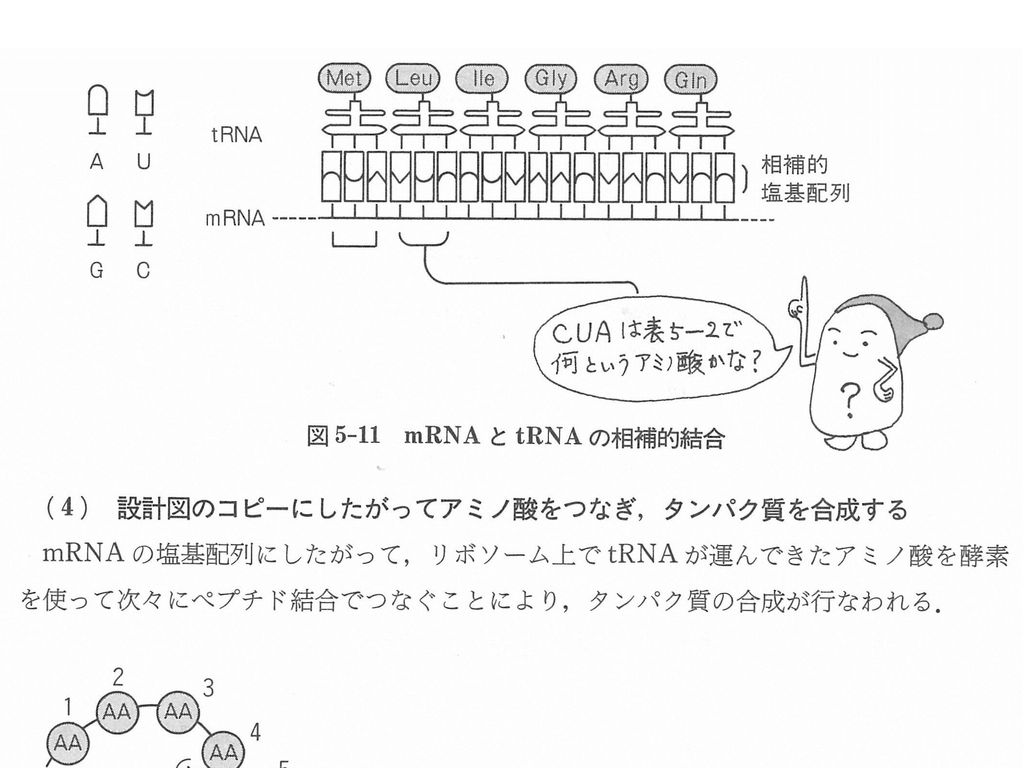
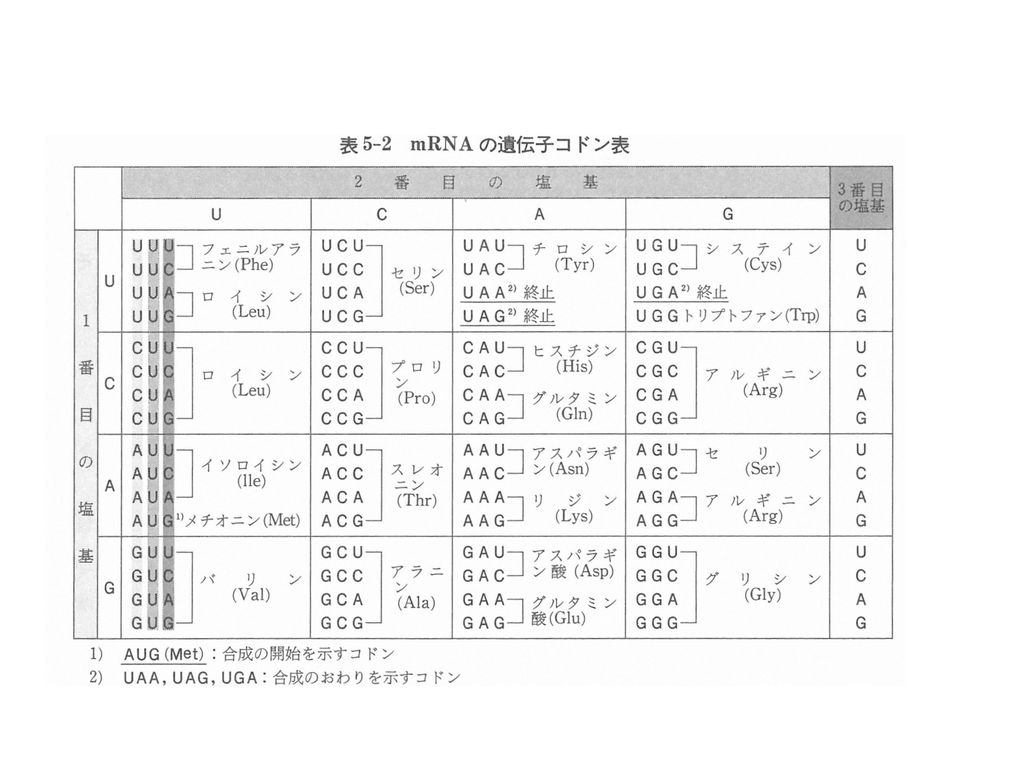
翻訳 Translation =タンパク合成 リボソーム上で mRNAの塩基配列がタンパク質のアミノ酸配列に変換される

29
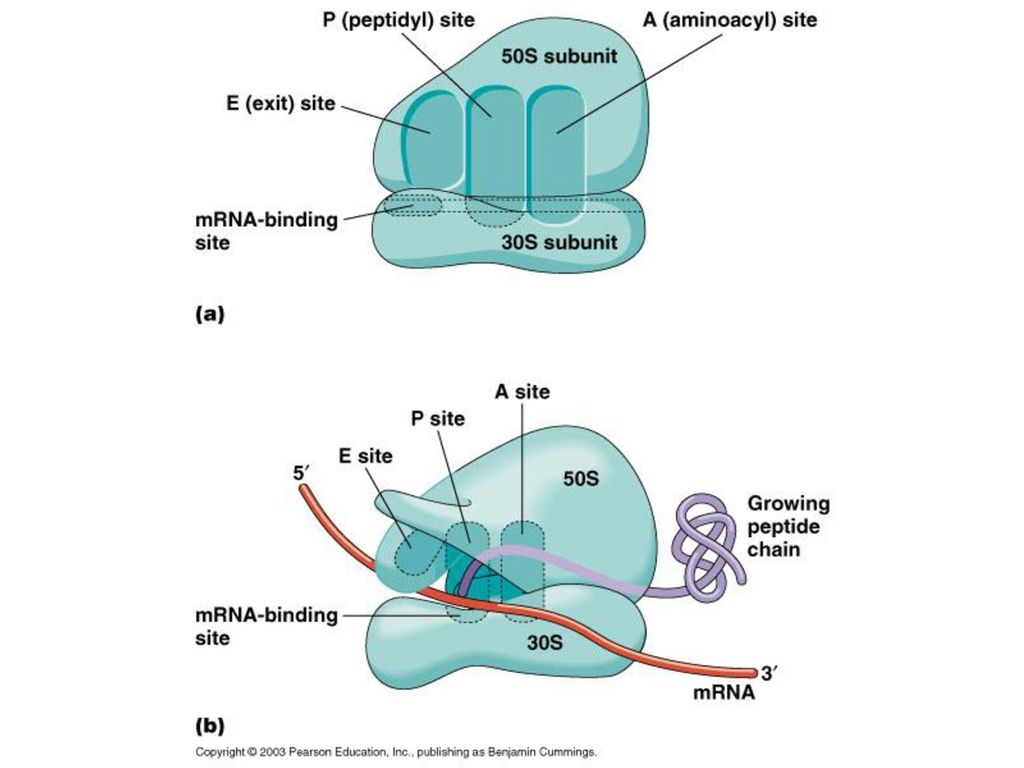
Ribosome 大サブユニット グレーとオレンジどちらがRNA, どちらがタンパク??

30
リボソーム 先の図オレンジがタンパク P-site-bound peptidyl tRNA, 緑;
SSU (40S) proteins, 青緑; small subunit (18S) rRNA, 黄色yellow LSU (60S) proteins, オレンジ; large subunit (25S/5.8S/5S) rRNAs, 青.
 proteins, 青緑; small subunit (18S) rRNA, 黄色yellow. LSU (60S) proteins, オレンジ; large subunit (25S/5.8S/5S) rRNAs, 青.")
31
小胞体(ER)膜についているPoly(ribo)some
膜についているPoly(ribo)some")
32
転写と翻訳 (バクテリアでは転写と翻訳は同時に起こる)
")
37
1. An aminoacyl-tRNA molecule binds to a vacant A-site on the ribosome 2. A new peptide bond is formed at the P-site (LSUずれる) 3. The mRNA moves a distance of three nucleotides through the small-subunit chain (SSUが移動) Figure Translating an mRNA molecule. Each amino acid added to the growing end of a polypeptide chain is selected by complementary base-pairing between the anticodon on its attached tRNA molecule and the next codon on the mRNA chain. Because only one of the many types of tRNA molecules in a cell can base-pair with each codon, the codon determines the specific amino acid to be added to the growing polypeptide chain. The three-step cycle shown is repeated over and over during the synthesis of a protein. An aminoacyl-tRNA molecule binds to a vacant A-site on the ribosome in step 1, a new peptide bond is formed in step 2, and the mRNA moves a distance of three nucleotides through the small-subunit chain in step 3, ejecting the spent tRNA molecule and “resetting” the ribosome so that the next aminoacyl-tRNA molecule can bind. Although the figure shows a large movement of the small ribosome subunit relative to the large subunit, the conformational changes that actually take place in the ribosome during translation are more subtle. It is likely that they involve a series of small rearrangements within each subunit as well as several small shifts between the two subunits. As indicated, the mRNA is translated in the 5′-to-3′ direction, and the N-terminal end of a protein is made first, with each cycle adding one amino acid to the C-terminus of the polypeptide chain. The position at which the growing peptide chain is attached to a tRNA does not change during the elongation cycle: it is always linked to the tRNA present in the P site of the large subunit.
 Figure Translating an mRNA molecule. Each amino acid added to the growing end of a polypeptide chain is selected by complementary base-pairing between the anticodon on its attached tRNA molecule and the next codon on the mRNA chain. Because only one of the many types of tRNA molecules in a cell can base-pair with each codon, the codon determines the specific amino acid to be added to the growing polypeptide chain. The three-step cycle shown is repeated over and over during the synthesis of a protein. An aminoacyl-tRNA molecule binds to a vacant A-site on the ribosome in step 1, a new peptide bond is formed in step 2, and the mRNA moves a distance of three nucleotides through the small-subunit chain in step 3, ejecting the spent tRNA molecule and resetting the ribosome so that the next aminoacyl-tRNA molecule can bind. Although the figure shows a large movement of the small ribosome subunit relative to the large subunit, the conformational changes that actually take place in the ribosome during translation are more subtle. It is likely that they involve a series of small rearrangements within each subunit as well as several small shifts between the two subunits. As indicated, the mRNA is translated in the 5′-to-3′ direction, and the N-terminal end of a protein is made first, with each cycle adding one amino acid to the C-terminus of the polypeptide chain. The position at which the growing peptide chain is attached to a tRNA does not change during the elongation cycle: it is always linked to the tRNA present in the P site of the large subunit.")
38
翻訳の素過程

39
系統樹を作るための予備知識 遺伝子の系譜≒種の系譜 なのだが 遺伝子の系譜を正確に求めるのは困難(化石がない) どんな遺伝子を比較するか
遺伝子の系譜≒種の系譜 なのだが 遺伝子の系譜を正確に求めるのは困難(化石がない) どんな遺伝子を比較するか 相同遺伝子を比較する 相同遺伝子には種類がある 小数の遺伝子をいい加減に比較したのでは、正確な系統樹は得られない computerを利用して大量の正確な情報を用いる
 どんな遺伝子を比較するか. 相同遺伝子を比較する 相同遺伝子には種類がある. 小数の遺伝子をいい加減に比較したのでは、正確な系統樹は得られない computerを利用して大量の正確な情報を用いる.")
40
パラログ、オルソログ、ホモログ パラロジー(paralogy): 1つの種の中で塩基配列がよく似ていること、種形成以前に進化的に近い関係であった。 パラログ(paralog):遺伝子重複で生まれた類似遺伝子で、同一ではないが、類似の機能を持っているタンパクの遺伝子。 オーソロジー(orthology): 異なる種の間で塩基配列がよく似ていること。 オルソログ(ortholog):共通の先祖遺伝子から垂直に伝えられ(通常の遺伝)、異なる種で同じ機能を示すタンパクの遺伝子。 ホモロジー(homology): 相同性、種の内部、種間でも共通の進化的起源を示唆する類似性。(収斂進化ということもある) ホモログ(homolog):相同性を示す配列をもったもの。
: 異なる種の間で塩基配列がよく似ていること。 オルソログ(ortholog):共通の先祖遺伝子から垂直に伝えられ(通常の遺伝)、異なる種で同じ機能を示すタンパクの遺伝子。 ホモロジー(homology): 相同性、種の内部、種間でも共通の進化的起源を示唆する類似性。(収斂進化ということもある) ホモログ(homolog):相同性を示す配列をもったもの。")
41
種の系譜(系統樹)と遺伝子の系譜
と遺伝子の系譜")
42
全生物の系統樹を作るのに適当な遺伝子 全生物が持っている生命に必須なもの rRNA(リボソームRNA) リボソームタンパク質
例 Ribosomal protein L1 ATP合成酵素と真核生物のV-ATPase

43
rRNAs

44
rRNA tree

45
ATPase

47
細胞質 リボソームが多数

48
Cell wall(外膜)と 細胞膜 鞭毛基部体とATP合成酵素-二つのロータリーモーター
と 細胞膜 鞭毛基部体とATP合成酵素-二つのロータリーモーター")
50
F-ATPase Movies

51
Fo部分

53
Gogarten, J.P. and Taiz, L. (1992) Photosynth. Res. 33: 137-146.
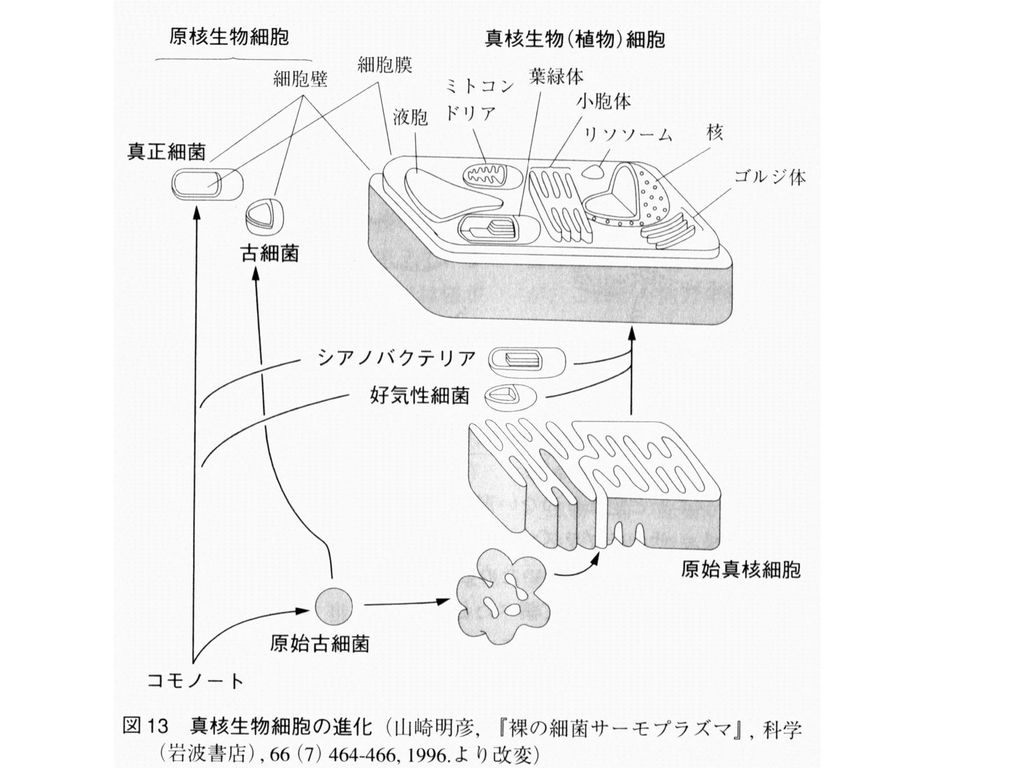
ATP合成酵素から液胞膜V-ATPaseが進化した (ATPを利用して液胞内部を酸性にする) 真核生物 V-ATPase 古細菌 真正細菌
 真核生物. V-ATPase. 古細菌. 真正細菌.")
54
外群 古細菌 菌類 ニンジン ハエ ラット 細菌 トウモロコシ葉緑体 ラン藻 菌類 ミトコンドリア トウモロコシ ミトコンドリア ラット ミトコンドリア 古細菌ATP合成酵素 真核生物液胞膜ATPase 真正細菌ATP合成酵素

55
現在でも遺伝子の移動は起こっている ミトコンドリア プラスチド 核 核には,ミトコンドリアゲノム丸ごとあるいは多数の断片
ミトコンドリア プラスチド 核には,ミトコンドリアゲノム丸ごとあるいは多数の断片 常にプラスチドゲノムから核への移動が起こっている なぜか,プラスチドゲノムには移動しにくい?? 核






 は塩基数が 200 ~ 400 程度と短い環状の一本鎖 RNA のみで構成 され、維管束植物に対して感染性を持つもの。分子内で塩基対を形成し、多くは 生体内で棒状の構造をとると考えられる。 ウイルスは蛋白質でできた殻で覆われているがウイロイドにはそれがなく、また プラスミドのようにそのゲノム上にタンパク質をコードすることもない。複製は.>")




 ・たてがき / 縦書き (from right to.>")







>")
.>")




